癫痫的治疗和药物发现现状
2021-02-17
小分子药物免疫疗法
▲速抢免费参会名额 | 大湾区(广州)生物医药创新者峰会
癫痫的治疗和药物发现现状

Preview
来源: 药融圈
来源
药学学报
作者
刘颖,于海波,孔庆飞
哈尔滨医科大学神经生物学教研室
中国医学科学院、北京协和医学院药物研究所,天然药物活性物质与功能国家重点实验室
摘要
关键词
_正文_
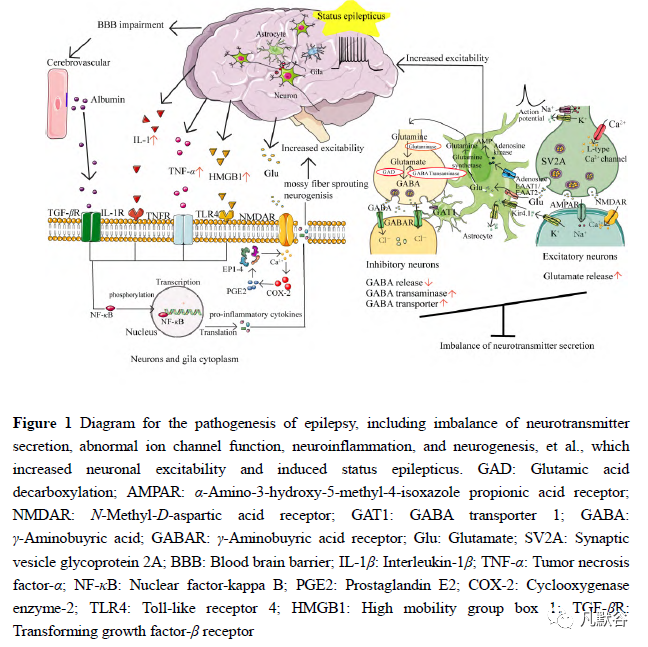
1癫痫的发病机制
Preview
来源: 药融圈
1.1 离子通道功能异常
离子通道是调节神经元兴奋性的基础,研究发现有多种离子通道基因与癫痫发生有关,主要包括钠离子通道、钾离子通道和钙离子通道等,编码这些通道的基因突变可影响离子通道正常生理功能,从而导致遗传性疾病[6]。
电压门控性钠通道(voltage gated sodium channels,VGSCs)主要负责动作电位的产生和传递,随着基因测序技术的进步,已经检测出4 种与癫痫相关的钠通道基因亚型变异,包括SCN1A、SCN2A、SCN3A 和SCN8A 基因,这4 个钠通道基因在抑制性和兴奋性神经元中皆有表达,但是它们表达强度各异。编码Nav1.1 的电压门控性钠通道SCN1A 主要存在于脑内γ-氨基丁酸(γ-aminobuyric,GABA)能抑制中间神经元的轴突起始段,等位基因功能丢失性突变可导致GABA 能抑制中间神经元放电功能障碍而引起严重的难治性癫痫,比如德拉韦综合征(Dravet syndrome,DS)[7]。而其余几个钠通道基因常发生功能获得性突变,导致神经元兴奋性异常升高。
电压门控性钾通道对神经元兴奋性调节至关重要,包括维持静息膜电位、调节动作电位放电以及释放神经递质等功能。研究表明,编码钾通道的基因变异会引起不同的癫痫发作。迄今为止,已发现多种癫痫相关的钾通道突变基因[6,8],如Kv1 通道基因LGI1、KCNA1 和KCNA2 功能丢失性突变,会导致不同类型的癫痫发作[8]。KCNT1 基因错义突变与常染色体显性遗传性夜间额叶癫痫(autosomal-dominant nocturnal frontal lobe epilepsy,ADNFLE)和婴儿期恶性迁移性灶性癫痫(malignant migrating focal seizures of infancy,MMFSI)有关[9]。KCNQ2 和KCNQ3 基因分别编码Kv7.2 和Kv7.3 通道,Kv7.2/Kv7.3 异构体组装形成的M 电流是神经元兴奋性的主要调节因子,M 电流具有稳定静息膜电位以及限制神经元重复放电的作用。实验证实,突变诱导的Kv7.2 亚基的稳定性降低或者M 电流门控的改变可能会导致新生儿癫痫的发生[10,11]。
Ca2+通过电压门控性钙通道进入细胞,作为第二信使启动细胞内事件,如收缩、分泌、突触传递和基因表达等[12]。电压依赖性钙通道是最早被发现与失神癫痫相关的离子通道。根据通道激活的膜电位范围,钙通道主要分为两种类型,高电压激活的钙通道(L 型、P/Q 型和N 型)以及低电压激活的钙通道(T 型钙通道)[12]。在癫痫领域,低压激活的T 型钙通道被研究的最多,包括3 种不同的亚型:Cav3.1、Cav3.2 和Cav3.3,分别由CACNA1G、CACNA1H 和CACNA1I基因编码[13]。目前,已明确钙离子通道致病性突变基因主要发生在CACNA1A(Cav2.1)和CACNA1H (Cav3.2) 两种基因,与之相关的疾病是儿童失神性癫痫和遗传性全面性癫痫。失神发作是皮质-丘脑-皮质环路异常所致,CACNA1A(Cav2.1)是失神癫痫的易感基因,通道异常会影响膜兴奋性及依赖钙内流的神经递质释放,环路发生异常震荡节律最终导致失神发作[14]。T 型钙通道基因表达异常,在动作电位过程中可以观察到显著的钙内流,导致神经元兴奋升高。如编码Cav3.2 的CACNA1H 基因突变,导致皮质-丘脑突触的兴奋性神经传递减少,但是,如何导致网络兴奋性增加的机制尚不清楚[6]。
1.2 中枢神经系统兴奋性/抑制性神经递质失衡
癫痫与中枢神经系统突触间兴奋性和抑制性神经递质的分泌不平衡有关,神经递质识别膜上的特定受体蛋白,这些蛋白将信息转化为细胞可以整合和传递的新信号。主要的神经递质涉及两大系统,促离子型受体 [也称为配体门控离子通道 (ligand-gated ion channel,LGIC)] 和代谢型受体 [G 蛋白偶联受体 (G proteincoupled receptor, GPCR)][15]。GABA 和谷氨酸分别作为中枢神经系统中最主要的抑制性和兴奋性神经递质,维持着膜的稳定性。
GABA 作用于神经元的A 型γ-氨基丁酸(type-Aγ-aminobuyric,GABAA)受体可引起快速的跨膜氯电流反应,使突触后膜产生超极化。研究发现,GABAA1 基因突变削弱了GABA 能抑制反应,导致癫痫发作[6]。离子型谷氨酸受体(ionotropic glutamate receptors, iGluRs)主要位于神经元的突触部位,当突触前释放的谷氨酸作用于突触后膜的iGluRs 时,可诱发突触的兴奋反应,这对神经元之间的通信及突触可塑性调节等过程具有极为重要的作用。根据iGluRs 对不同激动剂的相对选择性及其氨基酸序列的同源程度,可将其分为α-氨基-3-羟基-4-恶唑-丙酸受体(α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor, AMPA)、海人藻酸受体(kainite receptor, KA)和N-甲基-D-天冬氨酸受体(N-methyl-D-aspartate receptor,NMDA)[16]。AMPA 受体的GluA2 亚基组成的通道仅对Na+离子选择性通透,同时阻止Ca2+内流,AMPA 受体阻断剂可抑制癫痫的发生。相反,NMDA 受体主要对Ca2+通透,受体激活后会增加突触可塑性以及促进电点燃的诱导。同时,研究报道参与编码NMDA 受体的基因错义突变可导致早发型癫痫和智力障碍[17]。
1.3 神经炎症和胶质系统异常
癫痫发作后,脑神经元或活化的血管会释放炎症介质,进而促进炎症基因的自分泌和/或旁分泌的转录激活。值得注意的是,炎症介质也能直接激活表达在致痫组织神经元上的同源受体,后者诱导谷氨酸和GABA 受体及其他离子通道等的转录活性改变,使神经元兴奋性增加[20-22]。如胶质细胞和神经元上表达的Toll 样受体是一类固有免疫受体,在癫痫发作后或反复发作期间,内源性IL-1β和高迁移率族蛋白B1(high mobility group box 1,HMGB1)释放增多,它们作为TLR4(Toll-like receptor 4)的配体,与TLR4 结合后使下游的胞内信号分子活化,后者可以诱导核转录因子κB (nuclear factor-kappa B,NF-κB) 调节的炎症基因转录。
1.4 神经环路异常
癫痫的形成与大脑神经元间突触联系异常和病理性神经环路的建立而导致的神经网络兴奋性增强有关。近年研究发现,长时程增强(long term potentiation,LTP)、NF-κB 基因调控蛋白、苔藓纤维出芽 (mossy fiber sprouting, MFS)、神经肽Y (neuropeptide Y, NPY)、神经细胞黏附分子 (neural cell adhesion molecule,NCAM) 以及代谢型谷氨酸受体(metabotropic glutamate receptors,mGluRs)等多种因素均参与癫痫的发生发展和神经元突触的可塑性过程。在癫痫动物模型或者患者的海马组织中发现,癫痫发作可诱导致癫痫区的谷氨酸能突触产生LTP,并且该突触间联系发生持久性的重构[29]。NF-κB 存在于神经元胞体附近的突触末端,被活化后参与丝裂原活化蛋白激酶 (mitogen-activated protein kinases,MAPK) 信号通路,以调控神经元自身状态。
有研究者[30]提出在癫痫持续状态中,NF-κB 与其他转录因子共同作用,上调海马组织的转录活性,从而增加癫痫的易感性。MFS 是颞叶癫痫最主要的病理改变,多项研究表明,癫痫发作时,semaphorins[31]、ephrins[32]等多种轴突导向分子和MFS 表达异常。正常情况下,这些导向分子参与轴突的生长,与相应的膜受体结合后,引起膜内信号传导和细胞骨架重排,使轴突向正确的方向生长,然而,在病理状态下,轴突的生长方向受影响,最终导致异常神经元网络的形成和MFS 的改变。正常大脑的齿状回结构作为一种滤过器,阻滞癫痫发作从内嗅皮质向海马的传导,齿状回的这种滤过功能在某种程度上依赖于颗粒细胞之间近乎缺如的单突触联系[33]。在癫痫的动物模型中发现,MFS 会投射到齿状回颗粒细胞中谷氨酸能中间神经元上,导致齿状回兴奋性增加并传递给海马区[34]。
1.5 耐药性癫痫
在啮齿动物模型中,与AEDs 治疗有效的模型相比,观察到耐药性模型的脑组织中P-糖蛋白 (P-glycoprotein,P-gp) 高表达,而且发现P-gp抑制剂能减弱AEDs 的抗性[42]。但是,该假说仅在动物模型上被验证,尚不清楚其研究结果是否与难治性癫痫患者一致,同时,癫痫诱发的血脑屏障上P-gp 的表达上调是否影响脑内致痫区的药物浓度或者说P-gp 过表达是否只是癫痫发作的偶发现象仍存在争议。药物对靶点 (如离子通道或受体) 的反应性降低也是潜在的耐药机制,比如实验观察到卡马西平在耐药性癫痫中对钠通道的阻断作用减弱[43]。癫痫发作诱导的神经网络的退化和重构会抑制内源性癫痫发作系统,从而抑制AEDs 进入神经靶点,这也被认为与癫痫耐药性有关。
有研究观察到丙戊酸(valproic acid,VPA)在点燃大鼠模型中有不同反应,即对药物治疗有反应和对药物治疗有抗性。进一步研究发现,黑质网状带(substantia nigra pars reticulata,SNr)可能是导致癫痫耐药性的靶点,VPA 的抗癫痫作用可能是与SNr 神经元上的GABA 能神经元有关,并且它能调节SNr 的放电模型,使SNr 达到生理状态下的规律性[44]。另外,有研究者[45]提出在癫痫模型中选择性增强海马沟回锥体神经元兴奋性会增加对苯妥因的抗性,认为该区域的神经元异常活动与癫痫耐药性有关。然而,并没有实验证明在对药物治疗反应良好和具有抗性的癫痫患者之间是否存在神经网络异常,因此,需要更多的实验来验证这一假说。
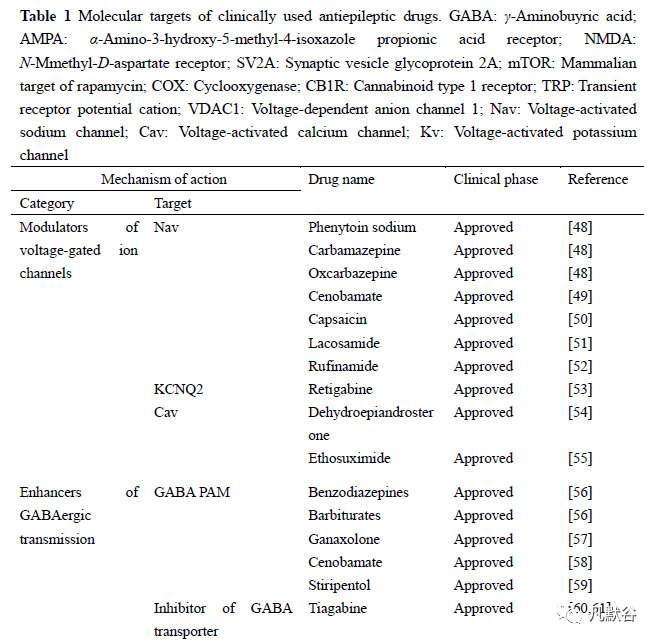
2现有治疗癫痫药物及作用机制
Preview
来源: 药融圈
Preview
来源: 药融圈
2.1 以电压门控离子通道为靶点的药物
2.1.1 以钠离子通道为靶点的药物
众所周知,SCN1A 基因突变会导致严重的癫痫发作,其中研究最多的是DS。DS 是一种儿童癫痫性脑病,在生命早期发病,随后会伴随认知和行为功能发育迟缓,最终导致突发性死亡[7,85],其发病机制与SCN1A 基因功能丢失性突变有关。因为DS 严重影响患者的生活质量,研究者一直致力于寻找更加安全有效的药物来治疗该病。研究证明,使用钠通道阻断剂会使患者病情恶化,而增强GABA能中间神经元传递的化合物则能有效治疗DS。同时,研究发现针对DS 患者最有效的方法是补偿Nav1.1 单倍体功能不足的精确疗法,其中,蜘蛛毒肽Hm1a能够选择性增强Nav1.1 的活性,进而恢复小鼠模型中抑制性中间神经元的功能[7,86]。
SCN2A 基因突变导致通道功能获得性突变,钠通道活性增加,易产生一系列癫痫综合征和新生儿癫痫的发作,该类癫痫对钠通道阻断剂 [苯妥英钠、卡马西平、LTG 和拉考沙胺] 有良好的反应[6,51]。SCN3A 基因突变与早期婴儿癫痫性脑病有关,与SCN2A 相似,通道功能获得性活性增加,该疾病对钠离子通道阻断剂有效[8]。SCN8A 基因功能获得性突变会引起严重的癫痫性脑病,新型钠通道调节剂GS967 对持续性钠电流升高有治疗效果,另一种钠通道抑制剂Prax330在SCN8A-N1768D 突变 [SCN8A(D/+)] 敲入小鼠中,可以降低持续性和复苏性钠电流,并且抑制动作电位爆发,降低神经元兴奋性[87,88]。除了与这4 种钠离子通道基因突变有关的癫痫发作类型,还有其他与钠通道异常有关的疾病。Lennox-Gastaut 综合征(Lennox-Gastaut syndrome,LGS)是一种严重的儿童期发育性癫痫性脑病,以耐药性癫痫、智力障碍和脑电图异常为特征,具有认知和运动障碍。VPA 是治疗LGS 的一线药物之一,卢非酰胺(rufinamide,RFN)主要限制钠离子依赖的动作电位,已被证实对LGS 相关的癫痫发作有效[52,89]。
2.1.2 以钾通道为靶点的药物
电压门控性钾通道是种类最多的离子通道,有大约40 余个基因编码,这些通道对神经元兴奋性调节至关重要[6,90]。迄今为止,仅有两种以钾通道为靶点的药物被证实具有抗癫痫作用,即氟哌啶和瑞替加宾,瑞替加宾是氟哌啶的结构类似物,这两种化合物均作用于Kv7.2/Kv7.3 钾通道[91]。作为Kv7.2/Kv7.3 的激动剂,主要稳定Kv7.2 的开放通道构象,并将激活曲线左移到更负的膜电位[91]。瑞替加宾已于2011 年被批准用于癫痫患者的辅助治疗药物。虽然氟哌啶对治疗新生儿癫痫有效,但是长期应用会引起药源性肝损伤,因此,不建议使用氟哌啶进行癫痫的治疗。而长期使用瑞替加宾的患者会出现色素沉着以及视力损伤等不良反应。因瑞替加宾的药理作用和不良反应的作用机制不同,故研究者一直致力于寻找既能稳定Kv7.2/Kv7.3 活性又能降低不良反应的化合物[92]。此外,研究发现通过钾通道以外的机制也可以缓解钾通道相关癫痫的发作,如阻断持续性钠通道和NMDA 受体拮抗剂均能有效治疗Kv1 通道有关的癫痫[93]。
2.1.3 以钙离子通道为靶点的药物
目前,关于高电压激活钙通道与癫痫发生有关系的报道较少,研究表明,神经类固醇脱氢表雄酮(dehydroepiandrosterone,DHEA)能调控CaV2.1α 亚型而发挥抗癫痫作用[54]。CaV2.3 通道抑制剂可能对癫痫有潜在的治疗效果,但目前仍缺乏选择性的小分子抑制剂[94]。
与高压激活钙通道不同,调节神经元T 型钙通道是非常有前景的癫痫治疗策略。实验证实,敲除CACNA1H 基因可以治疗T 型钙通道基因突变相关的癫痫[6]。乙琥胺是最早用于治疗失神癫痫的抗癫痫药,作为T 型钙通道的有机小分子阻断剂,该化合物对失活通道有较高的亲和力,可状态依赖性阻断T 型钙通道[55,95]。有文章报道以T 型钙通道为靶点,用来治疗全身性癫痫的化合物ACT-709478 正在进行II 期临床试验。另外,临床上应用的抗癫痫药物唑尼沙胺和丙戊酸也被证实对T 型钙通道有调节作用。虽然T 型钙通道阻断剂是治疗癫痫的潜在药物靶点,但是由于该通道广泛分布于多种组织中,且与其他电压门控通道同源性较近,对设计选择性T 型钙通道阻断剂挑战性非常大,因此还需突破这一限制,寻找更加有效的化合物[13]。
2.2 以神经突触为靶点的药物
2.2.1 调节GABA 能系统的药物
除了增强GABA 受体活性的药物,还有其他选择性作用于GABA 能系统的新型抗癫痫药物正在研究中,如噻加宾和维卡他林。前者通过选择性抑制γ 氨基丁酸转运体1(γ-aminobuyric acid transporter-1,GAT1)引起神经元和胶质细胞减少GABA 的摄取,后者通过抑制GABA 氨基转移酶而增加突触中GABA 的浓度。噻加宾开启了对GABA 转运体研究的进程,随后GABA 类似物EF1502(N-[4,4-bis(3-methyl-2-thienyl)-3-butenyl]-4-(methylamino)-4,5,6,7-tetrahydrobenzo[d]isoxazol-3-ol) 通过抑制GABA 转运体GAT1 显示出抗惊厥特性。加巴喷丁为GABA的前体,容易通透血脑屏障进入脑内,可以增加脑内突触GABA浓度[60,61]。噻吩戊醇(stiripentol,STP)作为辅助治疗药物,在临床治疗浓度下,通过与巴比妥盐相似的作用方式增强中枢GABA 传递[59]。西诺氨酯是一种新型抗癫痫药物,可作为高亲和力GABA 受体的正变构调节剂,不依赖苯二氮卓类药物结合位点而激活GABA 受体活性,并有效地增强海马神经元的抑制作用,这可能是稳定癫痫海马回路的潜在机制[58]。神经类固醇加奈索酮具有抗惊厥作用,其作用机制通过与GABAA 受体结合,增加氯通道通透性从而增强GABA 的神经元抑制作用[57]。
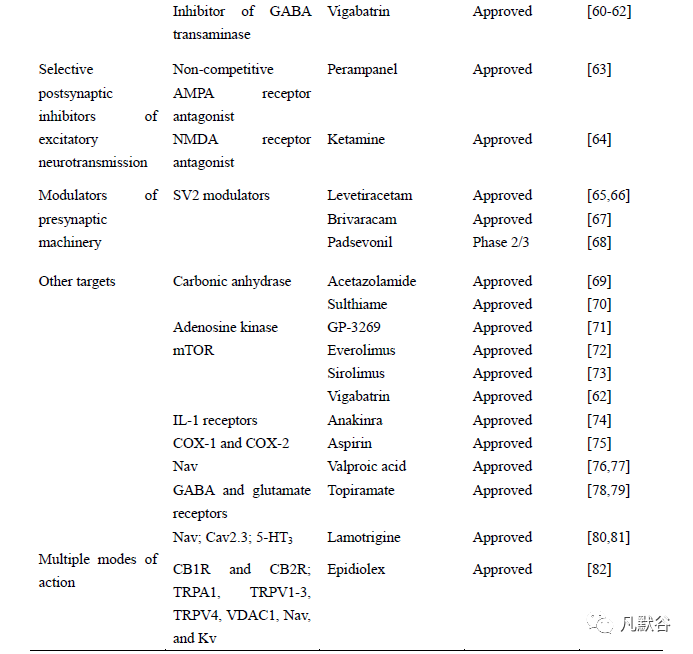
2.2.2 以谷氨酸受体为靶点的药物
谷氨酸是哺乳动物中枢神经系统内最主要的兴奋性神经递质,谷氨酸受体可分为离子型和代谢型两大类,前者属于LGIC,后者属于GPCR[97]。谷氨酸受体的表达和/或功能上调与癫痫发作有关[98]。AMPA 受体介导了大多数兴奋性神经递质传递,当AMPA 受体被阻断时,癫痫同步不能发生,而阻断KA 受体对癫痫的产生没有影响,NMDA 阻断剂只是部分消除癫痫样活动[99]。因此,AMPA受体已经成为引人注目的抗癫痫药物靶点,目前更多的关注在于开发更有效、耐受性更好的非竞争性AMPA 受体拮抗剂。有研究认为非竞争性拮抗剂通过充当跨膜片段之间的楔子来稳定AMPA 受体的关闭状态,从而抑制离子通道开放所必需的门控重排[100,101]。吡仑帕奈作为临床唯一被批准的非竞争性AMPA 受体拮抗剂,用于辅助治疗12 岁以上癫痫患者的部分发作和原发性全身性强直-阵挛发作[63] 。AMPA 受体的竞争性拮抗剂NBQX(2,3-dioxo-6-nitro-1,2,3,4tetrahydrobenzo[f]quinoxaline-7-sulfonamide),主要用于治疗难治性局灶性癫痫[102]。LTG 也能抑制AMPA 受体,减少谷氨酸释放[103]。此外,早期发现的几种NMDA 受体拮抗剂均能抑制谷氨酸释放,包括托吡酯(topiramate,TPM)和唑尼沙胺。NMDA 拮抗剂氯胺酮在癫痫持续状态动物模型中单独使用,没有显示出稳定的效果,但与其他药物联合使用却具有协同作用。尽管这几种NMDA 受体拮抗剂能够控制癫痫,但是目前尚无足够的证据证明其疗效和安全性[60,64]。
mGluRs 与G 蛋白偶联,一般将其分为3 组,第一组包括mGluR1 和mGluR5,第二组包括mGluR2 和mGluR3,第三组包括mGluR4、mGluR6、mGluR7 和mGluR8。多项研究显示,在颞叶癫痫患者和动物模型中第一组mGluRs 表达上调会增加神经网络的兴奋性,并且mGlu5 受体的负变构调节剂似乎是有望于治疗痉挛性癫痫综合征的候选药物。而第二和第三组mGluRs 对癫痫有抑制作用[17]。目前,市场上还没有发现以mGluRs 为靶点的抗癫痫药物。但是,研究者仍致力于这类药物的开发。因为mGluRs 的特性,针对mGluRs 的药物治疗可以在不干扰iGluRs 信号传递的基础上控制中枢神经系统谷氨酸能信号的获取,减少因阻断离子型谷氨酸受体而引起的不良反应[104]。
2.2.3 突触囊泡糖蛋白2(synaptic vesicle glycoprotein 2,SV2)
神经元间信号传递依赖突触前神经递质的释放,而这些神经递质的释放涉及数十种蛋白质。其中就包含SV2 家族。SV2 家族由3 个成员组成:SV2A、SV2B和SV2C。SV2 存在于分泌小泡(包括突触小泡),具有促进突触囊泡功能的作用[105]。纯合子SV2A 基因敲除小鼠会导致癫痫发生,甚至死亡[65]。左拉西坦作为一种新型抗癫痫药物,其特殊的作用机制就是与SV2A 结合位点结合,发挥抗癫痫作用。研究证明,突触囊泡中编码左拉西坦结合位点的SV2A基因变异与癫痫易感性有关。虽然不清楚左拉西坦与SV2A 结合后的具体机制,但是研究发现,SV2A 构象变化可能是其发挥癫痫保护作用的主要原因[65,66,106]。Brivaracam (BRV) 是一种选择性的高亲和力SV2A 配体,最近已被批准用于治疗难治性癫痫的部分发作[67]。抗癫痫候选药物padsevonil 同时作用于突触前SV2和突触后GABA 受体靶点,与地西泮相比,该药物的这种组合作用方式提供了较强的神经元保护作用[68,107]。这些药物的发现说明开发针对SV2A 靶点的抗癫痫药物具有无限潜能。
2.3 作用于其他靶点的药物
2.3.1 碳酸酐酶 (carbonic anhydrase,CA) 抑制剂
CA 是一种金属酶,目前共发现有15 种亚型,其中与神经元兴奋性有关的亚型包括VII、XII 和XIV[108]。CA 能催化CO2/HCO3-的可逆水合/脱水反应[109]。CA 在癫痫模型中活性较高,导致HCO3-内流增加,使突触后GABAA 受体去极化,从而启动和维持发作性放电[69]。因此,CA 抑制剂已成为治疗癫痫的一个间接途径。乙酰唑胺主要用于治疗女性月经性癫痫及部分和全身性癫痫发作。研究发现,乙酰唑胺可缩短发作时间和发作间隔,并降低GABA 能神经传递的持续时间和间隔,这些药理作用可能主要依赖于GABAA 受体介导的HCO3-生成减少,导致神经元去极化减弱,但因研究有限和不良反应严重,已经很少被使用[69,70,109]。而其他CA 抑制剂,如硫噻嗪也被证明能减少动作电位和癫痫样发作频率,唑尼沙胺用于成人部分癫痫的辅助治疗,而吲哚磺酰胺类药物也作用于CA靶点,表现出良好的抗惊厥活性[70,110]。TPM 是一种新型抗癫痫药,可用于治疗不同的癫痫类型,作用于多种靶点,其中就包括抑制血液及大脑中的CA,导致CO2 滞留而发挥抗癫痫的作用[108,109]。
2.3.2 腺苷激酶抑制剂
腺苷是一种内源性神经调节剂,能恢复损伤的神经组织,从而达到神经保护的作用[111]。腺苷的抗癫痫和神经保护作用主要是通过神经元和胶质细胞表达的腺苷A1 受体介导,与腺苷A1 受体结合,通过调节神经递质释放来影响神经元兴奋性,抑制突触前兴奋性神经递质释放,阻断突触后膜腺苷的感觉传递并引起膜电位超极化[112]。腺苷激酶是腺苷的上游调节因子,通过磷酸化AMP 来清除腺苷[113]。癫痫标志性的病理特征是星形胶质细胞增生,而后者通常与腺苷激酶的过度表达有关,腺苷激酶抑制剂被认为是能更有效提高突触腺苷水平的药物,因此,腺苷激酶是一种很有潜力的癫痫治疗靶点,其中GP-3269 是目前较为理想的候选药物[71]。
2.3.3 雷帕霉素靶蛋白 (mammalian target of rapamycin, mTOR) 抑制剂
结节性硬化症(tuberous sclerosis complex,TSC)最常见的神经系统症状是癫痫,通常在出生后第一年开始发病,同时还伴有神经发育不良以及认知问题[114]。TSC 是由TSC1 和TSC2 基因突变引起的,在正常情况下,TSC1 和TSC2整合生长信号和营养输入,使哺乳动物中mTOR 信号下调。mTOR 是一种进化上保守的丝氨酸-苏氨酸激酶,控制着细胞生长和存活[72]。研究发现,由于TSC基因突变,哺乳动物mTOR 信号通路的靶点过度激活引起巨细胞神经元细胞形态改变,异常的突触发生以及兴奋/抑制神经传递失衡[115]。因此,开发mTOR 抑制剂对治疗TSC 至关重要。依维莫司是选择性mTOR 抑制剂,具有良好的抗惊厥作用,其治疗安全性和耐受性良好,被美国食品药品监督管理局批准用于治疗TSC 伴发的癫痫发作[72,116]。西罗莫司与依维莫司的分子机制相似,对TSC 所致的癫痫发作也有显著的疗效[73]。
2.3.4 抗炎药
在神经炎症所致的获得性癫痫的发作过程中,发现在疾病早期抗炎药能有效治疗急性癫痫,并且抗炎药具有改善癫痫的神经并发症和神经保护作用。目前,临床上已经使用针对以下几种神经炎症通路为靶点的抗炎药来治疗癫痫。阿那白滞素是一种IL-1 受体拮抗剂,有报道称该药在治疗儿童癫痫综合征中,能减少癫痫发作次数和复发风险[74]。选择性COX-2 抑制剂,如罗非昔布和塞来昔布,能增加急性癫痫模型中癫痫发作阈值,但似乎不能用于治疗癫痫持续状态,而且研究证实,该类药需要合适的给药时间窗,否则将导致更严重的后果[25]。非选择性COX-1 和COX-2 抑制剂,如在匹罗卡品诱导的慢性癫痫模型中,阿司匹林能降低癫痫发作频率,具有神经保护作用且减少病灶中苔藓纤维出芽。而且局灶性癫痫患者在服用阿司匹林后,脑电图上显示减少发作频率[75]。最近发现多种以神经炎症通路为靶点的重组人源化单克隆抗体用于治疗癫痫患者,如阿达木单抗、托西珠单抗和那他珠单抗,它们分别以TNF、IL-6 和淋巴细胞为靶点,有效降低癫痫发作频率[117]。由此可知,若这些药物对预后不良或耐药性癫痫患者疗效显著,可以用抗炎疗法来干预治疗更多癫痫患者。
2.3.5 作用多种靶点的抗癫痫药物
2.3.5.1 VPA
2.3.5.2 TPM
2.3.5.3 LTG
2.3.5.4 大麻二酚(cannabidiol,CBD)
因此,认为CBD 的神经保护功能可能还与以下靶点有关,包括5-HT1A、CB1R、CB2R、TRP 通道家族(A1、V1~3 和V4)、电压依赖性阴离子通道1(voltage-dependent anion channel 1, VDAC1)以及钾通道等[127]。值得注意的是,表达在突触前的G 蛋白偶联受体55(GPR55)能调节突触前膜Ca2+释放,从而使神经元兴奋。研究发现,CBD 可通过拮抗GPR55 来调节神经元的兴奋性和突触可塑性[128]。目前,临床上具有抗癫痫作用的大麻素类药物主要是大麻二酚(商品名为Epidiolex),该药是美国食品药品监管局批准的首个含有大麻成分的药物,是一种高纯度的CBD 提取物,可用于LGS 和DS 的治疗[82]。
3药物发现现状
临床前筛选抗癫痫药物主要依赖于癫痫模型的建立,其中包括体外模型和体内模型。体外模型(动物脑切片和人体脑部异常放电组织等)是一种在体外条件下诱导脑组织癫痫样活动,并且模拟人类耐药性相关机制的模型,具有简化实验和节约资源的优点。然而,由于它们不能完全复制复杂神经元活动,通常还是选择体内模型来做药物筛查。目前,应用最广泛的是最大电休克(maximalelectroshock seizure,MES)和皮下戊四唑(subcutaneous pentylenetetrazole,SCPTZ)等急性癫痫模型。该类模型用于初步筛查具有抗惊厥活性的药物。然而,随着耐药性患者比重加大,研究发现这些急性癫痫模型不能检测治疗耐药性癫痫及慢性自发性癫痫的药物,因此需要其他实验模型来验证活性化合物在治疗耐药性方面的潜力[132]。随后便发现了病因相关模型,希望通过这些模型来进一步阐明癫痫的发病机制以及评估药物对癫痫患者的药理机制,它们分别为:
另外,精准医学治疗癫痫也是研究热点。靶向特殊脑区给药已经被证实比全身性给药更加有针对性。针对部分药效显著,但是不良反应严重以至于不能应用到临床上的药物,可以考虑靶向给药。靶向给药有以下几种策略,包括药物修饰、血脑屏障修饰和直接给药。药物修饰是基于聚合物和脂质的纳米载体,将药物输送到特定部位,增加脑内药物浓度。血脑屏障修饰可以通过抑制外排泵或者添加高渗性物质以改变血脑屏障的通透性,但是血脑屏障的损坏与癫痫有联系,不建议使用这种方法。直接给药是药物发挥作用最快捷的方式[136]。有研究在癫痫模型的黑质网状体上直接注射氨己烯酸,发现与腹腔注射相比,该药不仅增加癫痫发作阈值,而且相对减少外周的不良反应[62]。此外,靶向给药方法已经常规用于抗肿瘤治疗[137]。因此,靶向给药在癫痫治疗领域的广泛应用也指日可待。
4总结和展望
作者贡献:刘颖、于海波和孔庆飞共同参与了文章的撰写以及修改。
利益冲突:本文作者声明没有利益冲突。
参考文献
详见 药学学报
版权声明:本文转自凡默谷,如不希望被转载的媒体或个人可与我们联系,我们将立即删除
<END>
【关于药融圈】药融圈围绕我国生物医药产业链,针对生物医药大数据、技术和资本投资、药融园(产业园)等开展系列系统性工作,促进我国生物医药产业健康发展,完善产业链,共同面对全球合作和竞争。

Preview
来源: 药融圈

Preview
来源: 药融圈
点分享

Preview
来源: 药融圈
点点赞点在看
热门报告



立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。