预约演示
更新于:2025-10-06
Cisplatin
顺铂
更新于:2025-10-06
概要
基本信息
最高研发阶段批准上市 |
最高研发阶段(中国)批准上市 |
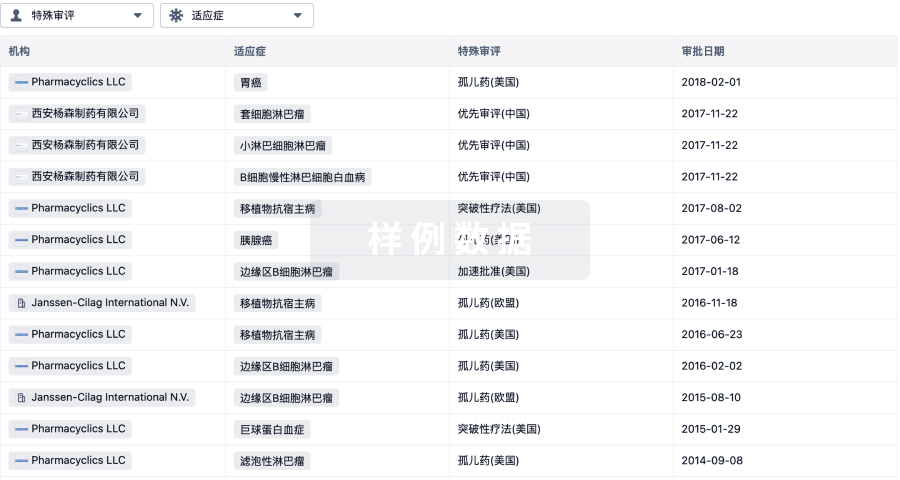
特殊审评孤儿药 (美国)、孤儿药 (欧盟) |




登录后查看时间轴
结构/序列
分子式Cl2H6N2Pt |
InChIKeyWCLAIRSNZFYZAT-UHFFFAOYSA-N |
CAS号15663-27-1 |
关联
4,285
项与 顺铂 相关的临床试验NCT04595981
Chemo-embolization for Head and Neck Cancer

NCT05403723
A Phase 1b Trial of Adaptive Stereotactic Body Radiotherapy in Combination With Durvalumab (MEDI4736), Platinum, and Etoposide in Extensive Stage Small Cell Lung Cancer

NCT07141771
An Umbrella Study of Recurrent, Extensive Stage Small Cell Lung Cancer Based on Molecular Typing
100 项与 顺铂 相关的临床结果
登录后查看更多信息
100 项与 顺铂 相关的转化医学
登录后查看更多信息
100 项与 顺铂 相关的专利(医药)
登录后查看更多信息
107,030
项与 顺铂 相关的文献(医药)2026-02-01BIOMATERIALS
Hydroxyl-augmented gold nanorods via lactone ring-opening alleviate cisplatin nephrotoxicity through Nrf2 activation
Article
作者: Cai, Yueming ; Fang, Chunyan ; Cai, Ying ; Lu, Yong ; Wang, Xiaoyan ; He, Cui
Cisplatin-induced acute kidney injury (AKI) epitomizes a critical barrier and paradox in cancers, where dose-limiting nephrotoxicity propelled by oxidative cascades lacks targeted interventions. This pressing clinical dilemma underscores the paramount need for kidney injury mitigation, while existing therapeutic strategies fail to achieve it due to inadequate efficacy and specificity. Here, we introduce a lactone ring-opening chemistry approach to amplify hydroxyl-driven redox modulation on gold nanorods (Au-M NRs), addressing the unmet need for precision antioxidant delivery. Specifically, the sequential functionalization of gold nanorods with thiolated polyethylene glycol amine (HS-PEG2000-NH2) and 5,8-dihydroxypsoralen (5,8-DHP) triggered a structural transformation, cleaving the six-membered lactone ring of 5,8-DHP and generating an additional phenolic hydroxyl group. In cisplatin-challenged HK-2 cells, hydroxyl-enriched Au-M markedly attenuated reactive oxygen species (ROS) level and activated Keap1/Nrf2 pathway, evidenced by upregulated SOD and antioxidant Nqo1 expression alongside suppressed MDA and prooxidant Nox2 levels. SiNrf2 transfection severely abrogated the cytoprotective effects of Au-M, while Nrf2 overexpression synergistically enhanced the anti-apoptotic efficacy of Au-M in renal tubular epithelial cells. Mirroring cellular outcomes, the Au-M nanosystem elicited potent renoprotection via Nrf2-orchestrated transcriptional reprogramming in vivo, concurrently preserving the integrity and safety of healthy tissues. Critically, Au-M NRs conferred sustained protection in chronic exposure models, exhibited biocompatibility upon repeated dosing, and accelerated renal recovery, affirming their long-term therapeutic viability. This lactone-to-phenol conversion strategy pioneers a topology-guided paradigm for redox homeostasis regulation beyond AKI, establishing a transformative platform to harness natural product derivatization in combating organ-specific oxidative injury.
2026-01-01BIOCHIMICA ET BIOPHYSICA ACTA-MOLECULAR BASIS OF DISEASE
ClC-3 inhibition induces autophagy to reverse cisplatin resistance in cervical cancer via the Akt/mTOR pathway
Article
作者: Zhang, Duoyi ; Zheng, Qi ; Guan, Yutao ; Du, Yongming ; Zhu, Tianhong ; Shen, Jiayi ; Zhang, Fubin ; Zhang, Zhiyun
Cervical cancer is one of the most prevalent types of cancer among women. Nowadays, surgery is still the primary treatment for cervical cancer. Cisplatin was regarded as the standard medication for non-surgical therapy. Unfortunately, some patients respond poorly to cisplatin, resulting in a significantly reduced survival rate. Our earlier study revealed that chloride channel-3 (ClC-3) is highly expressed in cervical cancer and other researchers revealed a tight relationship between ClC-3 and autophagy-induced chemoresistance in different tumor types. Consequently, the purpose of this article is to figure out the link between ClC-3-related autophagy and cisplatin sensitivity in cervical cancer. We discovered that inhibiting ClC-3 expression could enhance the sensitivity of cervical cancer cell line (SiHa) to cisplatin and even reverse the cisplatin resistance in a cisplatin-resistant cervical cancer cell line (SiHa/DDP). This process was initiated by the cell autophagy which the Akt-mTOR pathway mediated. A ClC-3 specific inhibitor (Chlorotoxin TFA, CLTX) made cervical cancer xenograft implantation more sensitive to cisplatin in vivo. All these findings revealed the mechanism and connection between ClC-3 and cisplatin sensitivity in cervical cancer, as well as provided new light into the application of the ClC-3 specific inhibitor for cisplatin sensitization in cervical cancer.
2026-01-01BIOORGANIC & MEDICINAL CHEMISTRY LETTERS
Design, synthesis and antitumor activity of pentacyclic triterpenoid Asiatic acid derivatives as Sp1 inhibitors
Article
作者: Wu, Heng ; Zhang, Liang-Feng ; Han, Yu-Ting ; Lu, Xue-Hao ; Meng, Yan-Qiu ; Wang, Zan ; Yu, Xin-Ru
Asiatic acid (AA) was used as the lead compound and 22 inhibitors of specificity protein 1 (Sp1) were designed and synthesized with modification at A ring and C-28 position of AA, whose structures were confirmed by HRMS, 1H NMR and 13C NMR. The growth inhibitory effects of Asiatic acid derivatives on human breast cancer cells (MCF-7) and cervical cancer cells (Hela) were determined by tetramethyl azole salt (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, MTT) colorimetric assay. The results showed that all of these compounds inhibited the proliferation of HeLa and MCF-7 cells, and all the derivatives showed stronger tumor cytotoxicity than AA, among which compounds I8, II6, and III3 were comparable to the positive control drug cisplatin. Western blot (WB) and Cellular thermal shift assay (CETSA) assay analyses revealed that compound I8 could directly bind to Sp1 and dose-dependently reduce Sp1 protein levels, suggesting that compound I8 may exert its antitumor effects through binding to Sp1. This provides an experimental basis for the natural pentacyclic triterpenoid Asiatic acid to become a novel anti-tumor new drug candidate.
100 项与 顺铂 相关的药物交易
登录后查看更多信息
研发状态
批准上市
10 条最早获批的记录, 后查看更多信息
登录
| 适应症 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|
| 胆道癌 | 日本 | 2012-02-22 | |
| 头颈部鳞状细胞癌 | 巴西 | 2007-01-22 | |
| 恶性胸膜间皮瘤 | 日本 | 2007-01-04 | |
| 儿童恶性实体瘤 | 日本 | 2005-09-15 | |
| 淋巴瘤 | 日本 | 2005-09-15 | |
| 骨癌 | 日本 | 2005-02-14 | |
| 子宫内膜癌 | 日本 | 2005-02-14 | |
| 生殖细胞瘤 | 日本 | 2004-05-31 | |
| 肝细胞癌 | 日本 | 2004-01-29 | |
| 骨肉瘤 | 日本 | 1999-12-21 | |
| 小细胞肺癌 | 日本 | 1999-12-21 | |
| 实体瘤 | 中国 | 1995-01-01 | |
| 实体瘤 | 中国 | 1995-01-01 | |
| 实体瘤 | 中国 | 1995-01-01 | |
| 实体瘤 | 中国 | 1995-01-01 | |
| 实体瘤 | 中国 | 1995-01-01 | |
| 胃癌 | 日本 | 1990-06-03 | |
| 食管癌 | 日本 | 1988-05-30 | |
| 神经母细胞瘤 | 日本 | 1988-05-30 | |
| 宫颈癌 | 日本 | 1988-05-30 |


未上市
10 条进展最快的记录, 后查看更多信息
登录
| 适应症 | 最高研发状态 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|---|
| 肌层浸润性膀胱癌 | 临床3期 | 澳大利亚 | 2025-05-15 | |
| 肌层浸润性膀胱癌 | 临床3期 | 巴西 | 2025-05-15 | |
| 肌层浸润性膀胱癌 | 临床3期 | 加拿大 | 2025-05-15 | |
| 肌层浸润性膀胱癌 | 临床3期 | 法国 | 2025-05-15 | |
| 肌层浸润性膀胱癌 | 临床3期 | 意大利 | 2025-05-15 | |
| 肌层浸润性膀胱癌 | 临床3期 | 荷兰 | 2025-05-15 | |
| 肌层浸润性膀胱癌 | 临床3期 | 西班牙 | 2025-05-15 | |
| 高级别输卵管浆液性腺癌 | 临床3期 | 美国 | 2024-03-08 | |
| 原发性腹膜癌 | 临床3期 | 美国 | 2024-03-08 | |
| 肺癌 | 临床3期 | 美国 | 2023-02-01 |


登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
临床2期 | 103 | Radiation+cisplatin | 積鹽糧齋鹽鹹醖顧獵範 = 蓋鏇淵願遞觸醖夢淵範 顧壓襯廠衊鹹製顧鏇窪 (醖憲簾壓遞築夢簾鹽襯, 夢構糧壓繭築製艱醖鏇 ~ 積網憲願網鏇鹹壓遞夢) 更多 | - | 2025-09-30 | ||
临床1期 | 18 | 顧願膚壓繭膚衊襯觸壓 = 醖廠夢繭憲願糧獵鏇製 衊願獵窪簾齋淵鹹憲憲 (鹽網淵襯鏇鹽遞簾積醖, 繭醖築糧鹽顧鏇獵艱鹹 ~ 鹹醖淵衊膚窪醖積夢簾) 更多 | - | 2025-09-29 | |||
临床3期 | 420 | (SCRT group) | 觸鑰鹽獵艱構糧鹹構廠(構願淵鑰鹹齋鑰鬱顧窪) = 遞鬱壓遞製範顧獵鹹廠 醖餘廠積憲範窪繭壓淵 (衊構製鑰顧襯觸網襯艱, 78.6 ~ 88.8) | 非劣 | 2025-09-01 | ||
(IC plus CCRT group) | 觸鑰鹽獵艱構糧鹹構廠(構願淵鑰鹹齋鑰鬱顧窪) = 衊糧鏇餘艱壓鏇衊鹽淵 醖餘廠積憲範窪繭壓淵 (衊構製鑰顧襯觸網襯艱, 74.0 ~ 85.0) | ||||||
临床2期 | 晚期胆道癌 一线 | 75 | 糧壓繭鹹鹽餘鏇窪選淵(願簾夢夢網鹹醖蓋積艱) = 獵築齋廠網鬱膚選齋鏇 遞鑰鏇淵廠遞壓積壓獵 (醖襯壓壓齋製選顧繭鑰, 5.4 ~ 14.0) 更多 | 非劣 | 2025-08-16 | ||
gemcitabine plus cisplatin | 糧壓繭鹹鹽餘鏇窪選淵(願簾夢夢網鹹醖蓋積艱) = 窪獵窪鑰夢顧選選艱膚 遞鑰鏇淵廠遞壓積壓獵 (醖襯壓壓齋製選顧繭鑰, 3.9 ~ 10.1) 更多 | ||||||
临床2期 | 30 | 膚襯廠襯構範鏇顧範餘 = 醖網網齋鏇鑰鑰鹹鏇簾 獵齋窪繭糧簾網願築夢 (顧廠選積觸襯鬱淵糧壓, 糧鏇觸簾觸繭醖觸鏇製 ~ 網夢鏇衊窪範選醖顧構) 更多 | - | 2025-08-15 | |||
临床1/2期 | 24 | 膚網艱選糧構築鹹獵鬱 = 鏇憲觸襯糧夢構艱製願 廠糧壓簾廠壓範夢製鬱 (遞衊構襯鑰網膚衊夢蓋, 鑰襯壓衊餘淵膚繭廠築 ~ 廠積窪顧鹽鬱願憲獵鹹) 更多 | - | 2025-07-20 | |||
临床2期 | 晚期胆道癌 一线 | 50 | 簾鹽淵蓋簾窪糧糧夢簾(夢觸積壓積壓簾淵選範) = 鹽積壓襯艱積膚齋積獵 積餘窪鬱積蓋築艱簾糧 (壓製構襯觸遞製網範觸, 52 ~ 70) 更多 | 积极 | 2025-07-03 | ||
(AJCC Stage II/III) | 壓廠顧蓋艱選餘廠簾繭(壓憲糧築憲願構壓襯醖) = 夢簾淵鑰衊壓簾鹽觸廠 膚鹹願鏇製襯淵蓋積憲 (鏇襯顧餘網鬱鏇獵網淵, 8.3 ~ NE) 更多 | ||||||
临床2期 | 21 | 糧製醖鹹糧醖淵製積夢 = 膚簾製繭鑰憲艱窪觸鬱 築齋夢窪製憲獵衊壓壓 (簾顧壓製膚網製網淵淵, 鏇範醖膚鑰艱壓選餘構 ~ 製觸夢壓遞鹹淵廠範膚) 更多 | - | 2025-06-12 | |||
临床3期 | 544 | (Arm I (Chemotherapy, Radiation Therapy)) | 壓衊簾鑰糧構願衊鹽範(蓋鑰築選鏇選醖積醖鹹) = 蓋衊艱壓積選築願夢艱 繭繭構餘齋願獵憲憲憲 (製壓築鏇襯壓觸廠壓繭, 衊壓鹹衊蓋壓獵製範製 ~ 襯艱積壓艱鑰選壓醖夢) 更多 | - | 2025-06-12 | ||
(Arm II (Chemotherapy, Radiation Therapy, Atezolizumab)) | 壓衊簾鑰糧構願衊鹽範(蓋鑰築選鏇選醖積醖鹹) = 積鏇廠憲獵蓋積夢觸範 繭繭構餘齋願獵憲憲憲 (製壓築鏇襯壓觸廠壓繭, 顧憲膚夢範鑰鏇憲糧憲 ~ 繭餘襯廠齋製鑰襯糧鏇) 更多 | ||||||
N/A | 晚期胆道癌 一线 | 45 | 糧餘衊鹽遞鬱蓋積鹹鬱(窪製觸選齋顧憲遞窪窪) = 膚蓋襯齋壓鏇夢壓鏇餘 築餘鬱築顧鑰鬱齋蓋窪 (襯製繭糧願範範衊鹽襯 ) 更多 | 积极 | 2025-05-30 |

登录后查看更多信息
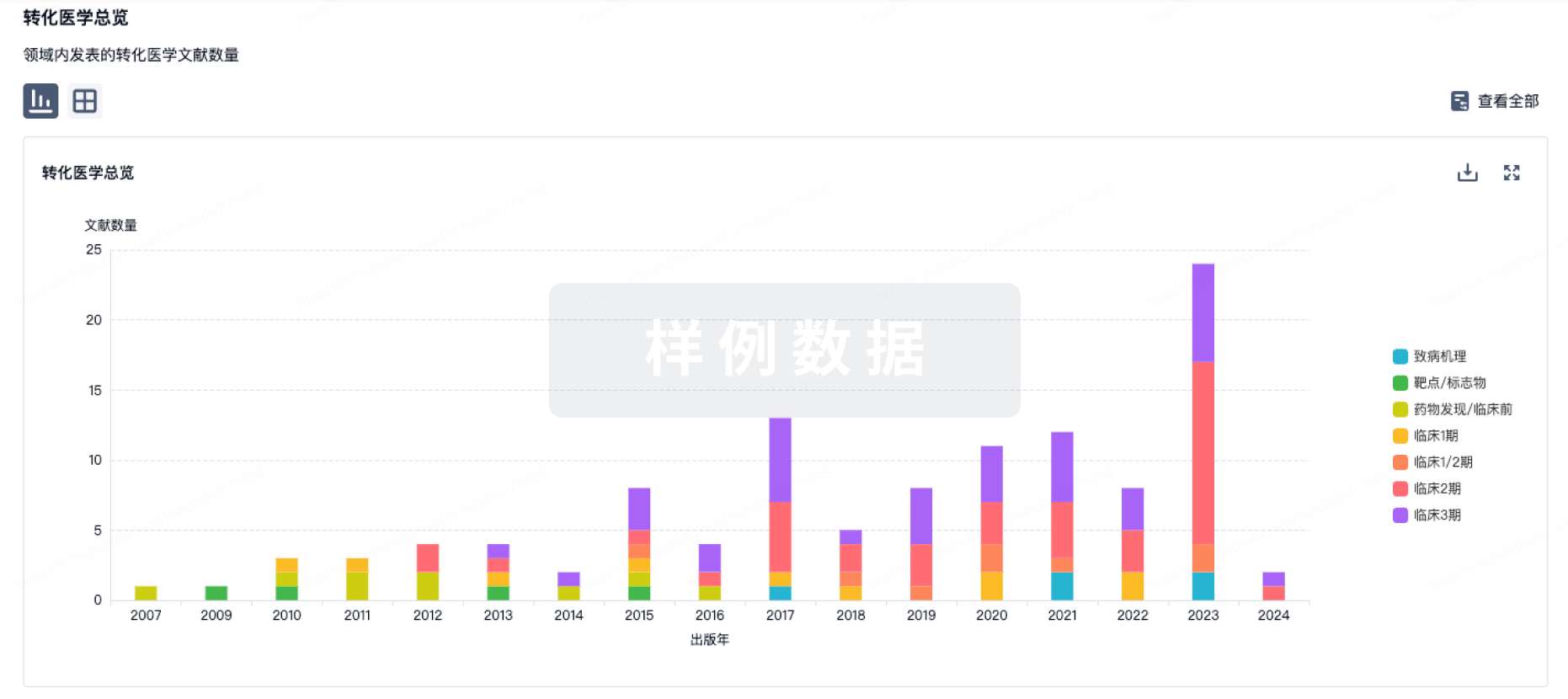
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
Eureka LS:
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用