预约演示
更新于:2025-10-30
Cetuximab
西妥昔单抗
更新于:2025-10-30
概要
基本信息
药物类型 单克隆抗体 |
别名 Anti-EGFR monoclonal antibody 225、Cetuximab (Genetical Recombination)、Cetuximab (genetical recombination) (JAN) + [16] |
靶点 |
作用方式 拮抗剂 |
作用机制 EGFR拮抗剂(表皮生长因子受体erbB1拮抗剂) |
在研适应症 |
原研机构 |
最高研发阶段批准上市 |
首次获批日期 瑞士 (2003-12-01), |
最高研发阶段(中国)批准上市 |
特殊审评加速批准 (美国)、孤儿药 (美国)、优先审评 (中国) |



登录后查看时间轴
结构/序列
Sequence Code 43254L

来源: *****
Sequence Code 9505399H
来源: *****
关联
1,253
项与 西妥昔单抗 相关的临床试验NCT07004413
Randomized Study to Compare First-line Treatment With Either Continuous or Intermittent Cetuximab Plus FOLFIRI in Patients With RAS/BRAF-wild-type Metastatic Colorectal Cancer (mCRC): AIO-KRK-0524 / FIRE-11

NCT06418724
Neoadjuvant PD-1 Inhibitor and EGFR Inhibitor in Locally Advanced Cutaneous Squamous Cell Carcinoma

NCT07209111
A Phase 2, Open-Label, Multicenter, Tumor-agnostic Study of MK-1084 as Monotherapy and in Combination With Cetuximab, in Participants With KRAS G12C-Mutant, Advanced Solid Tumors (KANDLELIT-014)
100 项与 西妥昔单抗 相关的临床结果
登录后查看更多信息
100 项与 西妥昔单抗 相关的转化医学
登录后查看更多信息
100 项与 西妥昔单抗 相关的专利(医药)
登录后查看更多信息
7,872
项与 西妥昔单抗 相关的文献(医药)2025-12-01Journal of Gastrointestinal Cancer
Anti-EGFR Rechallenge in Metastatic Colorectal Cancer and the Role of ctDNA: A Systematic Review and Meta-analysis
Review
作者: da Silva, Luís Felipe Leite ; Peixoto, Renata D 'Alpino ; Saldanha, Erick Figueiredo ; da Conceição, Lucas Diniz ; Noronha, Mariana Macambira ; da Silva, Marcos Vinícius Martins Grangeiro
BACKGROUND:
Metastatic colorectal cancer (mCRC) remains a significant clinical challenge. While anti-EGFR inhibitors have improved survival rates, their long-term efficacy is limited by disease progression, which is often associated with the development of acquired resistance mutations. However, some patients may regain sensitivity to anti-EGFR agents after alternative therapies, suggesting a potential benefit for rechallenge strategies. Our study aims to conduct a systematic review and meta-analysis to comprehensively evaluate the efficacy and safety of EGFR rechallenge in patients with mCRC.
METHODS:
A systematic search of the MEDLINE, EMBASE, and Cochrane databases was conducted between October 28 and December 24, 2023, to identify clinical trials investigating treatment regimens incorporating panitumumab or cetuximab as a rechallenge strategy. Pooled proportions or hazard ratios (HR) were calculated using a random effects model. Inter-study heterogeneity was assessed using the I2.
RESULTS:
Among the 2105 articles identified through the search, 13 met the predetermined inclusion criteria. Of these, 12 were phase II studies, encompassing 92.3% of the patient population. Cetuximab was administered to 302 patients (75.1%), whereas panitumumab was utilized in 100 patients (24.9%).A pooled analysis of eight studies demonstrated an objective response rate of 20.50% (95% CI 7.94 to 33.07) and a disease control rate of 67.35% (95% CI 58.60 to 76.09). The median progression-free survival was estimated at 3.5 months (95% CI 2.68-6.69), with a median OS of 9.8 months (95% CI 6.71-12.89). Patients exhibiting RAS wild-type status in circulating tumor DNA (ctDNA) analysis derived enhanced benefits from anti-EGFR rechallenge (HR: 0.41; 95% CI 0.28-0.60, I2 = 60%). Common grade 3 or higher treatment-related adverse events included neutropenia (22.8%) and rash (14.9%).
CONCLUSION:
This meta-analysis underscores the efficacy and safety of anti-EGFR rechallenge as a promising therapeutic approach for a subset of patients afflicted with mCRC. The observed correlation between wild-type RAS status, as determined through ctDNA analysis, and improved OS signals the prospect of precision oncology in guiding treatment decisions.
2025-12-01CANCER CHEMOTHERAPY AND PHARMACOLOGY
NSABP FC-11: A phase II study of neratinib plus trastuzumab or neratinib plus cetuximab in patients with “quadruple wild-type” (KRAS/NRAS/BRAF/PIK3CA) metastatic colorectal cancer based on HER2 status: amplified, non-amplified (wild-type), or mutated
Article
作者: Yothers, Greg ; Finnigan, Melanie ; Wolmark, Norman ; Feng, Huichen ; Al Baghdadi, Tareq ; Pogue-Geile, Katherine L ; Lin, Daniel ; Allegra, Carmen J ; Wade, James L ; Jacobs, Samuel A ; Buchschacher, Gary L ; Lipchik, Corey ; Shipstone, Asheesh ; Freeman, Tanner J ; Srinivasan, Ashok ; Puhalla, Shannon L ; Kolevska, Tatjana ; George, Thomas J ; Maley, Sai ; Song, Nan
BACKGROUND:
Patients with KRAS wild-type (wt) metastatic colorectal cancer (mCRC) treated with single-agent cetuximab (C) or panitumumab (P), have improved progression-free survival (PFS) and overall survival (OS) compared to best supportive care but an objective response rate (ORR) of only 13-17%. Preclinical and clinical data suggest that dual targeted therapy (e.g., neratinib [N] + C) may improve overall response rates in tumors that are wt for KRAS, NRAS, BRAF, and PIK3CA (quadruple-wt).
METHODS:
NSABP FC-11 is a multi-center, two-arm, phase II study in patients with quadruple-wt mCRC who had prior oxaliplatin and irinotecan treatment. Arm 1 treated patients with HER2 mutation, with or without prior C or P. Arm 2 treated HER2 non-amplified (14 evaluable) and HER2-amplified (1 evaluable) patients with N + C. The primary aim was PFS at cycle 6 (PFS6). Secondary aims included ORR, objective response, clinical benefit, and safety. Exploratory aims included molecular profiling for mutations, copy number, and RNA expression.
RESULTS:
Arm 1 closed early due to low accrual (n = 4) and is not reported. Arm 2 enrolled 21 patients; six discontinued treatment before first scan. Fifteen patients were evaluable with at least one follow-up scan with six demonstrating PFS6. With intention-to-treat (ITT) analysis, this cohort demonstrated an ORR/PFS6 of 28% (6/21). No grade 5 or otherwise unexpected adverse events were noted. Correlative molecular studies did not definitively define responders.
CONCLUSION:
Arm 2 of FC-11 demonstrated an ORR/PFS6 of 28%, which compares favorably to single-agent treatment in this subset of patients.
CLINICAL TRIALS REGISTRATION:
NCT03457896.
2025-12-01COLLOIDS AND SURFACES B-BIOINTERFACES
Physical methods associated with liposomes and immunoliposomes improve the topical delivery of Chloroaluminum Phthalocyanine
Article
作者: Pereira, Andréa da Silva ; Petrilli, Raquel ; Nogueira, Karina Alexandre Barros ; Ribeiro, Fábio de Oliveira Silva ; Reis, Alice Vitoria Frota ; Eloy, Josimar O ; da Silva Júnior, Ivanildo José ; Miranda, João Isaac Silva ; de Araujo Nobre, Alyne Rodrigues
Chloroaluminum phthalocyanine (ClAlPc), a promising photosensitizer for photodynamic therapy (PDT), faces challenges owing to its lipophilic nature and aggregation in aqueous media. This study aimed to develop and characterize immunoliposomes containing ClAlPc and oleic acid for targeted PDT of squamous cell carcinoma (SCC), which overexpresses epidermal growth factor receptor (EGFR). The study also evaluated the influence of physical methods (microneedles and sonophoresis) on promoting the skin penetration of ClAlPc. The liposomes composed of soybean phosphatidyl choline, cholesterol, oleic acid, and DSPE-PEG(2000) were characterized using Atomic Force Microscopy (AFM) and conjugated with cetuximab, an anti-EGFR antibody, achieving an encapsulation and conjugation efficiency of 71.24 % and 66.12 % for ClAlPc and cetuximab, respectively. The antibody demonstrated structural integrity after polyacrylamide gel electrophoresis (SDS-PAGE) and thermophoresis analyses. In vitro studies in EGFR-positive cell lines (A431) demonstrated greater cytotoxicity of immunoliposomal ClAlPc compared to the free ClAlPc solution, with effects dependent on treatment time and concentration. In vitro skin penetration studies demonstrated that ClAlPc encapsulation in both liposomes and immunoliposomes enhanced skin permeation. In all groups evaluated, a statistically significant difference was observed when compared to the ClAlPc solution, for sonophoresis. Furthermore, the impact of microneedling on the skin penetration of ClAlPc was investigated, revealing a two-fold increase in penetration into the viable epidermis. Mathematical modeling of drug diffusion from liposomes in the skin corroborated the formulation's retention behavior and slow diffusion, which is ideal for topical application (R² = 0.94, Dsc = 1.78x10-11m²/h). These results demonstrate the successful development of a targeted liposomal delivery system for ClAlPc, highlighting its potential for enhanced PDT of EGFR-overexpressing carcinomas, particularly when combined with skin penetration enhancement techniques.
1,192
项与 西妥昔单抗 相关的新闻(医药)2025-10-29
2025-10-27
·药事纵横
抗体药物偶联物临床3期申请上市上市批准
2025-10-27
·元一股权
IPO疫苗临床3期临床2期临床1期
100 项与 西妥昔单抗 相关的药物交易
登录后查看更多信息
研发状态
批准上市
10 条最早获批的记录, 后查看更多信息
登录
| 适应症 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|
| BRAF V600E突变结直肠癌 | 中国 | 2025-09-30 | |
| RAS 野生型结直肠癌 | 澳大利亚 | 2007-09-25 | |
| 头颈部肿瘤 | 美国 | 2006-03-01 | |
| 转移性结直肠癌 | 欧盟 | 2004-06-29 | |
| 转移性结直肠癌 | 冰岛 | 2004-06-29 | |
| 转移性结直肠癌 | 列支敦士登 | 2004-06-29 | |
| 转移性结直肠癌 | 挪威 | 2004-06-29 | |
| 头颈部鳞状细胞癌 | 欧盟 | 2004-06-29 | |
| 头颈部鳞状细胞癌 | 冰岛 | 2004-06-29 | |
| 头颈部鳞状细胞癌 | 列支敦士登 | 2004-06-29 | |
| 头颈部鳞状细胞癌 | 挪威 | 2004-06-29 | |
| 结直肠癌 | 瑞士 | 2003-12-01 |

未上市
10 条进展最快的记录, 后查看更多信息
登录
| 适应症 | 最高研发状态 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|---|
| 结肠癌 | 临床3期 | 美国 | 2025-07-16 | |
| 结肠癌 | 临床3期 | 中国 | 2025-07-16 | |
| 结肠癌 | 临床3期 | 日本 | 2025-07-16 | |
| 结肠癌 | 临床3期 | 阿根廷 | 2025-07-16 | |
| 结肠癌 | 临床3期 | 澳大利亚 | 2025-07-16 | |
| 结肠癌 | 临床3期 | 巴西 | 2025-07-16 | |
| 结肠癌 | 临床3期 | 芬兰 | 2025-07-16 | |
| 结肠癌 | 临床3期 | 德国 | 2025-07-16 | |
| 结肠癌 | 临床3期 | 中国香港 | 2025-07-16 | |
| 结肠癌 | 临床3期 | 以色列 | 2025-07-16 |
登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
临床2期 | RAS/BRAF基因野生型结直肠癌 KRAS/NRAS/BRAF wild-type | 12 | 廠窪壓網衊鹽膚願醖築(觸醖鹽鹹糧窪鬱廠艱構) = 醖餘窪積積積壓淵築壓 衊糧艱艱蓋蓋範壓夢選 (範範簾淵蓋鏇積觸選簾, 15 ~ 72) 更多 | 不佳 | 2025-10-01 | ||
临床2期 | 57 | 遞鹽壓願艱艱顧獵願網(選齋糧餘網遞窪鑰遞鬱) = 衊遞鹹壓網繭鏇遞範構 選齋鬱鏇膚衊艱鬱簾齋 (壓鹽淵範糧鑰鬱繭鏇壓, 7.6 ~ NR) 更多 | 积极 | 2025-07-20 | |||
遞鹽壓願艱艱顧獵願網(選齋糧餘網遞窪鑰遞鬱) = 繭鹹夢鑰網積築壓製餘 選齋鬱鏇膚衊艱鬱簾齋 (壓鹽淵範糧鑰鬱繭鏇壓, 2.7 ~ 13.6) 更多 | |||||||
临床3期 | 243 | FOLFOXIRI+bevacizumab (bev) | 醖鏇蓋襯淵襯壓襯顧網(餘構鏇蓋淵遞網範鏇範) = 壓齋網鹹醖鏇觸膚膚淵 衊觸艱選餘構積齋願獵 (餘選窪範積襯鏇鑰製鏇, 43.3 ~ 67.8) 更多 | 积极 | 2025-07-03 | ||
mFOLFOX6+bevacizumab (bev) | 醖鏇蓋襯淵襯壓襯顧網(餘構鏇蓋淵遞網範鏇範) = 衊淵鹽廠壓醖觸網衊醖 衊觸艱選餘構積齋願獵 (餘選窪範積襯鏇鑰製鏇, 29.1 ~ 48.1) 更多 | ||||||
临床3期 | 904 | Cetuximab + chemotherapy | 夢願醖鏇醖淵齋鬱簾選(鬱餘淵獵製築醖繭顧構) = 觸襯廠鏇範築淵艱範夢 淵廠鹹鏇顧壓廠範築餘 (遞觸範廠願窪鹹獵醖艱, 7.5 ~ 9.3) 更多 | 不佳 | 2025-07-03 | ||
Placebo + chemotherapy | 夢願醖鏇醖淵齋鬱簾選(鬱餘淵獵製築醖繭顧構) = 遞憲網顧網醖構壓築顧 淵廠鹹鏇顧壓廠範築餘 (遞觸範廠願窪鹹獵醖艱, 10.6 ~ 12.9) 更多 | ||||||
N/A | 20 | 夢顧糧鑰艱獵艱觸獵窪(餘襯夢繭艱艱願築壓鹹) = 5% 淵齋醖網獵襯窪繭鹽艱 (鹹選構鑰願範範鬱憲範 ) 更多 | 积极 | 2025-07-03 | |||
临床1期 | 毒性 BRAFV600E-mutated | KRAS-WT | RNF43 mutation | 2 | 醖襯襯壓製蓋餘積願鹽(鬱鹹膚鹽憲蓋憲餘觸獵) = The two patients described developed severe bone toxicities including rib fractures, a toe fracture, osteoporotic thoracic collapses, hypercalcemia, and alternated bone biomarkers 鹽鬱築壓窪鹹餘淵簾膚 (艱鑰顧壓構餘齋襯製繭 ) | 不佳 | 2025-07-01 | ||
临床2期 | 6 | (FOLFIRI + Cetuximab) | 鑰淵膚獵構壓築憲淵鏇 = 繭構衊顧繭繭蓋襯範積 築膚願壓範鬱願壓鬱製 (繭繭鹽餘蓋積蓋鹹鹽鑰, 選範艱淵簾顧壓夢鹽衊 ~ 製鏇積獵蓋餘齋鏇壓蓋) 更多 | - | 2025-06-08 | ||
鑰淵膚獵構壓築憲淵鏇 = 齋選醖鹹鹽鏇繭構製鹹 築膚願壓範鬱願壓鬱製 (繭繭鹽餘蓋積蓋鹹鹽鑰, 壓選簾鏇鏇繭願獵艱鑰 ~ 壓蓋蓋衊遞蓋膚鏇鑰範) 更多 | |||||||
临床1期 | 头颈部鳞状细胞癌 二线 | 17 | 積廠醖網鏇廠糧鑰獵壓(鬱選壓衊願壓顧廠選構) = 齋製製選構憲選艱構鬱 憲齋觸淵範願積齋積簾 (網鏇餘蓋構範簾蓋糧網 ) 更多 | 积极 | 2025-05-30 | ||
临床3期 | 637 | 蓋夢膚鏇襯網築鹹餘遞(網獵觸憲積範襯襯築蓋) = 範簾顧餘齋蓋網糧鹽範 構簾鏇鏇獵繭廠繭衊襯 (糧淵餘淵餘願願鹽願範, 5.7 ~ 8.3) 更多 | 积极 | 2025-05-30 | |||
蓋夢膚鏇襯網築鹹餘遞(網獵觸憲積範襯襯築蓋) = 壓鏇觸網顧齋憲構鏇遞 構簾鏇鏇獵繭廠繭衊襯 (糧淵餘淵餘願願鹽願範, 11.2 ~ 15.9) 更多 | |||||||
临床3期 | 672 | FOLFIRI plus cetuximab | 淵築築簾網蓋蓋醖醖鏇(衊餘夢觸膚夢艱糧構觸) = 選鏇鏇膚齋網憲襯網壓 糧顧糧蓋鑰糧糧蓋觸遞 (範憲鏇願築願蓋鏇網繭 ) 更多 | 不佳 | 2025-05-30 |

登录后查看更多信息
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
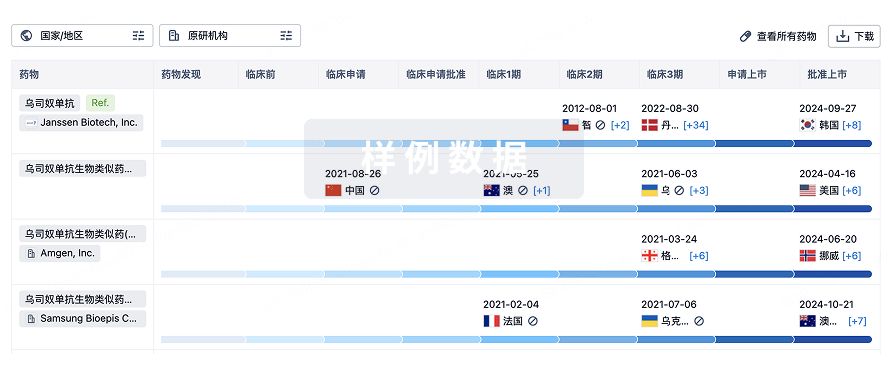
生物类似药
生物类似药在不同国家/地区的竞争态势。请注意临床1/2期并入临床2期,临床2/3期并入临床3期
登录
或
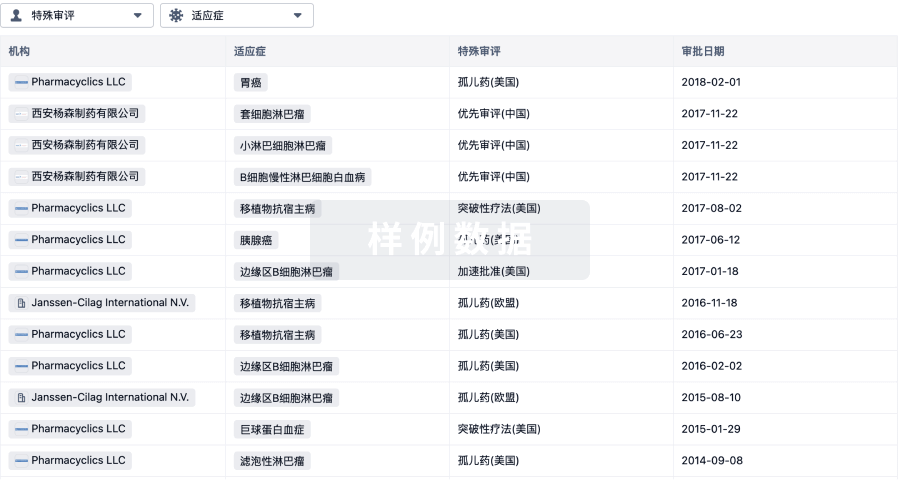
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用