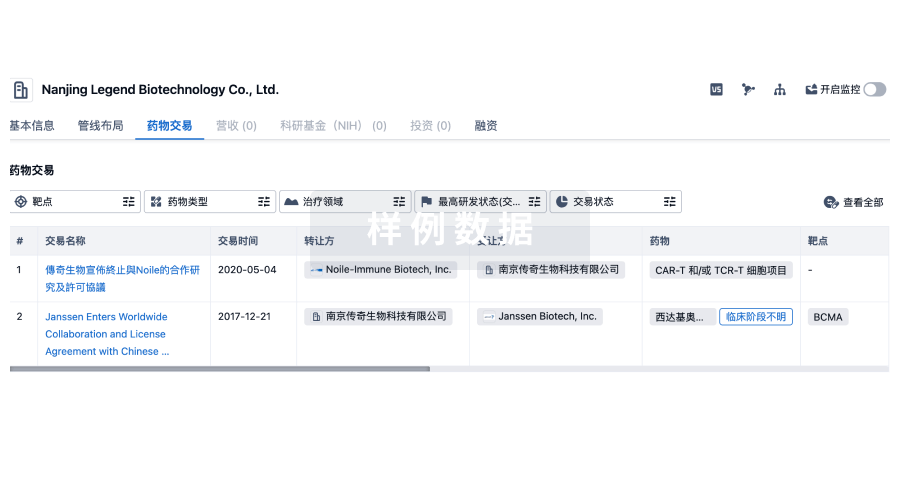
同写意年度巨献!!7月25-26日,邀您相聚金鸡湖畔,与5000人一起为中国医药创新“同写意”!1、双特异性或双靶点药物的崛起应和了如今药物研发的最大困境:小分子药和蛋白药的药物靶点不够用了。2006年时统计的已获批药物共针对324个药靶。2017年,已获批药物的药靶数量扩展到893个,其中667个是人蛋白。假设传统药物(不包括核酸药和细胞治疗)可开发的药靶共有1000个,所有生物技术和制药公司的共同竞争空间则受到极大的禁锢。但如果考虑双靶点药物的话,药靶空间(双药靶的随机组合数)增长了500倍。图片来源:参考资料132、双特异性抗体(这里简称双抗或bsAb)是最重要的一类双靶点药物,也是近些年来最热门的药物研发领域之一。它的研发历史已有几十年: 图中黄色的立柱代表和双抗有关的论文每年发表篇数(左轴),紫色的立柱代表每年进入临床试验的双抗数。到目前为止,已有三个双抗药获批,其中两个还在市场上(Amgen的Blincyto / Blinatumomab和Chugai的Hemlibra / Emicizumab),但有上百个双抗药分子正在临床试验中被评估。未来几年将是双抗药全面爆发的时代。3、双特异性抗体不等同于两种单克隆抗体的联合用药。和联合用药相比,双抗的优势包括单一的药代动力学特性(PK profile)和相对简单的申报流程:如果两种单抗在体内有不同的半衰期,在联合用药中需要平衡两者的给药时间表;联合用药在申报时需要准备两套CMC、毒理数据等。但双抗药最重要的优势是obligate effects,或全新功能。图片来源:参考资料144、如果一个双抗药的效果仅仅等同于(additive effects)或超过(synergistic effects)两种母单抗的联合用药产生的效果,它还不是最理想的双抗药。一个理想的双抗药可以产生全新的功能、全新的效果。如果这种新功能是两种母抗体不管如何联用也无法获得的,它就是obligate effects 。图片来源:参考资料185、双抗产生新功能主要有两个机理。第一个机理是通过空间重排产生新的药效(spatial obligate mechanisms)。在这种情况下,双抗同时结合两个药靶,把它们拉近,进而启动下一步行为。最常见的双抗——细胞桥接双抗——就是利用这个机理(a)。比如,T细胞导向双抗同时结合T细胞上的CD3和肿瘤细胞表面上的某种抗原(tumor associated antigen, TAA),桥接两种细胞,并激活T细胞,以杀死肿瘤细胞。还有的双抗,如b和c所示,同时结合同一细胞表面上的两种受体,或者导致细胞凋亡,或者促进细胞生长。这种效果只有在两种受体被交联时才会发生。Hemlibra则是d图描绘的双抗类型的一个例子:Hemlibra模仿FVIIIa因子,把FIXa和FX拉到一起,激活FX,引起血凝反应。图片来源:参考资料186、双抗产生新功能的第二个主要机理是在时间维度上完成的(temporal obligate mechanisms)。双抗分子先通过一个结合臂结合一个靶点,再因此被置换到新的空间(比如通过血脑屏障(图e),或被内吞入细胞(图f)),最后通过另一个结合臂结合第二个靶点,触发下游反应。如果第一个结合事件不发生,该双抗接触不到第二个靶点。从这个角度看,该双抗的第一个结合臂是第二个的递送装置。图片来源:参考资料147、图1中的499,500种双靶点组合只是可能的成药空间(双抗的成药空间比这要小的多)。理性设计只能找到寥寥不多的几种双抗。要从中找到更多的能产生新功能的组合需要一个高通量的筛选手段。对于双抗药来说,第一个难题是如何构建足够大的双抗分子库以供筛选。假设针对每个靶点有10种结合片段(binders)可供选择(覆盖不同的表位和亲和力),那么针对两个靶点就至少需要10 x 10 = 100种双抗分子。即使对大公司来说,制备、提纯100种蛋白分子也不是一件容易的事。这还只是在双靶点的成药空间中筛选。如果要从多靶点空间中寻找合适的双抗,制备的蛋白数量急剧增加。可以说,构建大规模的双抗分子库是双抗药研发的一个主要门槛。这里,我们以UCB的技术平台为代表来探讨一下制药公司是如何攻克这一难关的。在筛选阶段,UCB的所有结合片段都是以单臂Fab形式存在,这些Fab又分两种格式:Fab-Y和Fab-X。Fab-Y的C端带着一段多肽Y,而Fab-X的C端则带有可结合Y的scFv,两者混合后会形成一个临时的、可供筛选的Fab-KD-Fab双抗。Fab-KD-Fab技术的最大优势是节省了蛋白制备、纯化的成本。针对两种抗原A和B,UCB团队只需制备10种针对A的Fab-Y和10种针对B的Fab-X,一共20个蛋白,而不是100个蛋白。所有100种双抗都可以通过Fab-Y和Fab-X混合而生成。图片来源:参考资料148、UCB有一个项目是筛选抑制纤维化的双抗药物。人的肺(或肾)成纤维细胞与气管(或近端肾小管)上皮细胞混合培养,在培养液中加入不同的双抗分子,最后用免疫荧光方法比较各类蛋白纤维和细胞外基质的沉积程度。抑制纤维化可能的靶点有22个,双靶点的组合数就达到228种(理论上应该是231种可能:22 x 21 / 2), 再考虑针对同一靶点有多种Fab,一共有2000个双抗需要被制备、筛选。而UCB采用Fab-KD-Fab技术,只需制备130种Fab分子作为起始原料就可以覆盖所有可能的双抗。图片来源:参考资料149、 结果显示,一种双抗组合可以成功抑制细胞外基质的沉积或纤维化,而这种效果是两种单抗或其联合用药无法达到的。至此,UCB通过高通量筛选成功地找到一个抑制纤维化的候选双抗药物。其它公司也有自己的独特技术来构建大规模的双抗分子库,比如Genmab的DuoBody技术平台。10、一旦双靶点和双binder组合选定,下一步就要决定该双抗采取何种格式。双抗主要有三种格式:片段格式(左图),非对称的IgG类抗体格式(中图),和对称的IgG类抗体格式(右图)。每种格式里还有多种不同版本,图中给出的只是有代表性的例子。图片来源:Harpoontx.com/science/11、和IgG类双抗药相比,片段双抗主要有两个优势:由于其分子量小,它们可以更容易地渗透到肿瘤或靶向组织内,还可以结合到全长抗体不易接触的表位。但片段双抗的主要缺点是由于缺乏Fc,它们不能结合FcRn受体,因此体内半衰期短。比如Blincyto采取的是BiTE格式(左图),在体内的半衰期只有4个小时,其每个治疗周期需要连续28天静脉输液,给患者带来极大的不便,也增加了治疗费用。片段双抗可以通过改造而克服这一缺点。比如,Harpoon Therapeutics的TriTAC技术在片段双抗之间再加上能结合白蛋白(HSA)的抗体片段(右图)。片段双抗也可以和Fc融合而延长半衰期。图片来源:参考资料1812. 截至2019年6月份的数据表明,已进入或即将进入临床阶段的双抗药物中,非对称的IgG类双抗药占大部分——约60%。非对称IgG类双抗药的最大局限性是其生产过程中的效率问题。13、一个单克隆抗体是由两个相同的重链和两个相同的轻链构成。轻链与重链之间、重链与重链之间通过二硫键联结。在生产单抗蛋白时,先需要将两种质粒转染进同一个细胞(CHO或HEK293)里。一个质粒携带重链基因,另一个携带轻链基因。细胞同时表达重链蛋白和轻链蛋白,两种蛋白再自动组装成一个抗体分子,被细胞分泌到培养液里,不会有相似的副产物。但用细胞表达双抗分子时,事情就变得复杂一些。一个非对称IgG类双抗分子包含4个不同的蛋白链。宿主细胞需要被转染4种质粒,分泌出的抗体蛋白并不是100%想要的双抗。14、在细胞内,轻链与重链之间,重链与重链之间会随机配对。最后组装出的抗体分子会出现16种可能,10种蛋白分子。假设每种配对的概率都相同(实际上概率有些差异),我们所需要的双抗蛋白分子(红圈标出的)只占所有产物的1/8或12.5%。这不但导致了低效率的生产流程,而且还使提纯过程变得异常复杂:双抗很难从其它相似的抗体产物中分离出来。15、为了解决错配问题,工业界已经开发出很多创新技术。解决重链错配的常见技术有Genentech发明的Knobs-into-holes(KiH),Novimmune发明的Common Heavy Chain策略,和Regeneron发明的Fc双突变技术。KiH技术是在两种重链上引入不同的突变,使得同种重链无法配对,而异种重链则榫卯相扣。Common Heavy Chain策略则是使用同一个重链,对不同抗原的特异性主要由不同的轻链决定。Fc双突变技术与前两个技术不同。它不会导致单一配对,但便于选择性地提纯。一个重链还是野生型,另一个重链则引入两个突变(H435R和Y436F)。杂合重链双聚体与蛋白A的结合力比纯和突变重链双聚体高,但比纯和野生重链双聚体低。这样在蛋白提纯的过程中可以选择性地把含有杂合重链双聚体的双抗分子纯化出来。图片来源:参考资料1716、解决重轻链错配的常见技术有Common Light Chain策略,罗氏开发的CrossMab技术,Merck KGaA开发的CH1: CL界面改造技术,和药明生物、Invenra等公司开发的CH1: CL置换技术。Common Light Chain策略是使用同一个轻链,对不同抗原的特异性主要由不同的重链可变区决定。CrossMab技术(之一)是将一侧的轻重链的CH1和CL互相调换,这样野生的重链只能和野生的轻链配对,而含CL域的重链只能和含CH1的轻链配对。在CH1: CL界面改造技术中,一侧的CH1和CL各引入一系列突变,使得改造后的重、轻链依然能结合,但和野生型的轻、重链相排斥。药明生物的WuXiBody技术是通过将其中一对轻、重链的CH1/CL结构域置换为TCR Cα / Cβ而实现;与此相似,Invenra的B-body技术是将其中一对轻、重链的CH1/CL结构域置换为一对儿其它抗体结构域(未披露具体信息)。图片来源:参考资料13和1817、现在处于临床阶段的双抗候选药的85%是用来治疗肿瘤的,而抗肿瘤双抗药的大部分是T细胞导向双抗(T-cell redirecting bispecific antibodies, 简称TRBA),即有CD3结合臂的双抗。在剩下的篇幅中,让我们来重点讨论一下TRBA。图片来源:参考资料1118、T细胞表面的CD3和TCR组成CD3-TCR复合体,用来结合抗原呈递细胞(APC)表面的MHC-抗原片段。TRBA则绕开了MHC的限制,激活T细胞,把T细胞的火力引向了肿瘤细胞。TRBA的一个结合臂靶向肿瘤细胞表面的抗原(TAA),另一个结合臂结合T细胞表面的CD3ε,起到CD3ε激动剂的作用。TAA-TRBA-CD3-TCR形成了类似于MHC-peptide-TCR-CD3的免疫突触(IS),T细胞被激活,释放颗粒酶、穿孔素等蛋白,从而引起肿瘤细胞的裂解。图片来源:参考资料2419、正如前文所言,处于临床阶段的TRBA候选药物有很多种。Genentech的Mosunetuzumab就是其中之一。Mosunetuzumab是一个靶向B细胞表面的CD20的非对称全长双抗。它的Fc部分引进了一个突变(N297G), 去除了ADCC作用,以避免误伤T细胞。图片来源:www.biooncology.com/pipeline-molecules/cd20-tcb.html; 参考资料920、罗氏正在开发的Glofitamab(CD20-TCB或RG6026)是另一个TRBA例子。Glofitamab的独特之处在于它与CD20的结合是二价的,与CD3是单价的。这种格式增强了它杀死肿瘤细胞的能力。另外,它的Fc部分只保留了FcRn结合位点,但不会引起免疫效应,从而既保留了长的生物半衰期又不会误伤T细胞。从Glofitamab的结构示意图(右上角)可以看出,为了避免错配,这个分子的设计至少采用了三个技术:KiH,CrossMab和CH1:CL电荷界面。图片来源:参考资料2421、 T细胞导向双抗经常被拿来和另一种合成免疫(synthetic immunity)治疗手段——CAR-T作比较。作为第二代TRBA的代表,Mosunetuzumab、CD20TCB和Regeneron的REGN1979在一期临床试验中显示了对B淋巴瘤的良好疗效,不亚于已上市的CAR-T疗法。有两点需要说明的是,Kymriah(Tisagenlecleucel)和Yescarta (Axicabtagene Ciloleucel)的靶向抗原是CD19,而这三个TRBA靶向CD20。另外,TRBA和CAR-T之间还从没做有过头对头的临床试验比较。根据表中的数据作横向比较还需谨慎。ORR为客观应答率,CR为完全缓解率。图片来源:参考资料2422、CAR-T最常见的副作用是细胞因子风暴(CRS)和神经毒性。和已上市的CAR-T疗法相比,三个TRBA的这两项副作用的发生频率和严重程度都有降低。图片来源:参考资料1223、尽管如此,TRBA在药效和安全性上还有很大的提高空间。提高药效大致可以从7个方面下手:(A)T细胞结合臂靶向的受体不仅仅局限于CD3(由黄色的椭圆代表),还可以从CD28,CD2和αβTCR中选择;(B)T细胞结合臂可以靶向CD3的不同表位;(C)TRBA可以采取多种格式,包括片段双抗和类IgG双抗;(D)癌细胞表面上的TAA密度也影响TRBA的效果;(E)形成的TAA-TRBA-CD3-TCR的空间尺度对癌细胞的杀伤力也有影响,而这是由TRBA的格式、TAA的大小、TAA上的结合表位与癌细胞膜之间的距离等因素决定的;(F)增加TRBA与TAA的亲和力会增加对癌细胞的杀伤力;(G)调整TRBA与CD3的亲和力可改变TRBA的药效和毒性。(G)比较复杂。一般来说,TRBA与CD3的亲和力(affinity)或总亲和力(avidity)不能太高,否则将导致TRBA和循环中的T细胞率先结合,不能有效渗入肿瘤,也产生很多副作用。图片来源:参考资料624、让我们先看一个通过(A)来提高对肿瘤细胞杀伤力的例子。激活T细胞并使之持续增殖、保持对肿瘤细胞的杀伤力需要两个信号:第一个信号是由CD3-TCR复合体介导的,引起细胞因子的分泌;第二个信号是由CD28介导的,抑制T细胞凋亡。设计TRBA时一般只考虑了第一个信号,因此TRBA经常引起T细胞无能化(anergy)而限制了T细胞的持久杀伤作用。但双信号这个概念已经被应用在第二代、第三代CAR-T的设计中:CAR的膜内部分既包括CD3ζ(信号1),又包括CD28的膜内激活域(信号2)。图片来源:参考资料2025、Sanofi设计的三特异性抗体就同时靶向这两个信号通路。这个三抗的一个结合臂靶向骨髓瘤细胞表面上的CD38,另一个结合臂同时靶向CD3和CD28。为了降低引起CRS的风险,这个分子被设计成与CD3ε的亲和力相对不高(KD ~ 20 nM,而anti-CD28和anti-CD38与各自靶点的KD分别为2 nM和4 nM)。在细胞和动物实验中,CD38 / CD3 x CD28三抗显示了对肿瘤细胞的持久杀伤力。图片来源:参考资料2426、TRBA的主要副作用是CRS。有没有可能只保留TRBA对肿瘤细胞的裂解能力,但又剔除其引起CRS的风险?研究表明,细胞杀伤力和CRS都依赖于TCR刺激,但是它们需要不同强度的CD3结合水平。细胞毒性T细胞(CTL)有两种不同的激活阈值,引起细胞因子释放的CD3结合率阈值要比细胞裂解阈值高得多,至少相差百倍。因此,可以通过减弱其对T细胞的亲和力或增加其TAA / CD3结合价比例来设计更安全的TRBA。IGM Biosceinces设计的IGM2323就是这样一个例子。它由靶向CD20的IgM五聚体和一个靶向CD3的scFv构成。也就是说,IGM-2323对CD20和CD3的结合价之比为10:1。图片来源:参考资料2327、在细胞实验中,IgM版本的双抗比IgG版本显示了对肿瘤细胞的更强的杀伤力。图片来源:参考资料2328. 而同时在细胞和动物(包括猴子)实验中,和IgG版本双抗相比,IGM-2323几乎不引起细胞因子的释放。IGM-2323正在一期临床试验中被评估。图片来源:参考资料1729、增加TRBA安全性的另一个办法是,让它只在肿瘤微环境(TME)中才被激活,才能招募T细胞。Maverick Therapeutics的COBRA技术平台就是建立在这一机理上。MVC-101是通过COBRA推出的一个三抗候选药。它可以结合EGFR和HSA(延长生物半衰期),但只能有条件地结合CD3。在其它组织中,它结合CD3的VH和VL分别被VLi和VHi所屏蔽,不能结合CD3。肿瘤及其周边中富含MMP2和MMP9等基质金属蛋白酶。在MMP的作用下,分子内的衔接链被剪断,含VLi、VHi和anti-HSA的一半蛋白分子被释放。剩下的半个分子两两配对,形成了可四价结合EGFR、两价结合CD3的双聚体双抗。在取得了良好的细胞和动物数据后,MVC-101即将进入临床试验。图片来源:www.affimed.com/rock-platform/innate-cell-engagers/; 参考资料2630、招募T细胞不是桥接双抗的唯一策略。Affimed公司通过ROCK技术推出了一系列招募NK细胞的候选双抗药。ROCK双抗既可以结合肿瘤细胞表面的TAA,又可结合NK细胞表面的CD16A(即FcγRIIIa),通过激活先天免疫系统和ADCC来裂解肿瘤细胞。这种双抗既保留了杀伤肿瘤的药效,又降低了TRBA的CRS风险。Affimed的管线领头项目AFM13靶向CD30和T细胞淋巴瘤,目前正在二期临床试验中被评估。31、未来10年将是多特异性抗体药的爆发期。双抗药发展到今天,是几十年来学术界和工业界在概念上、技术上不断产生突破日积月累的结果。几十家生物技术和制药公司都做出了重要贡献。我在这里仅列出文中涉及的公司。在不久的未来,双抗药将被广泛用来治愈顽疾,拯救生命。到那时,我们不应忘记,它们的背后是一系列的创新和众多的开拓者。参考资料:(上下滑动查看更多)1. Overington, J. P., Al-Lazikani, B. & Hopkins, A. L. (2006) How many drug targets are there?, Nature reviews Drug discovery. 5, 993-6.2. Krah, S., Kolmar, H., Becker, S. & Zielonka, S. (2018) Engineering IgG-Like Bispecific Antibodies-An Overview, Antibodies (Basel). 7.3. Santos, R., Ursu, O., Gaulton, A., Bento, A. P., Donadi, R. S., Bologa, C. G., Karlsson, A., Al-Lazikani, B., Hersey, A., Oprea, T. I. & Overington, J. P. (2017) A comprehensive map of molecular drug targets, Nature reviews Drug discovery. 16, 19-34.4. Tustian, A. D., Endicott, C., Adams, B., Mattila, J. & Bak, H. (2016) Development of purification processes for fully human bispecific antibodies based upon modification of protein A binding avidity, MAbs. 8, 828-38.5. Klein, C., Schaefer, W., Regula, J. T., Dumontet, C., Brinkmann, U., Bacac, M. & Umana, P. (2019) Engineering therapeutic bispecific antibodies using CrossMab technology, Methods. 154, 21-31.6. Weinkove, R., George, P., Dasyam, N. & McLellan, A. D. (2019) Selecting costimulatory domains for chimeric antigen receptors: functional and clinical considerations, Clin Transl Immunology. 8, e1049.7. De Nardis, C., Hendriks, L. J. A., Poirier, E., Arvinte, T., Gros, P., Bakker, A. B. H. & de Kruif, J. (2017) A new approach for generating bispecific antibodies based on a common light chain format and the stable architecture of human immunoglobulin G1, J Biol Chem. 292, 14706-14717.8. Fischer, N., Elson, G., Magistrelli, G., Dheilly, E., Fouque, N., Laurendon, A., Gueneau, F., Ravn, U., Depoisier, J. F., Moine, V., Raimondi, S., Malinge, P., Di Grazia, L., Rousseau, F., Poitevin, Y., Calloud, S., Cayatte, P. A., Alcoz, M., Pontini, G., Fagete, S., Broyer, L., Corbier, M., Schrag, D., Didelot, G., Bosson, N., Costes, N., Cons, L., Buatois, V., Johnson, Z., Ferlin, W., Masternak, K. & Kosco-Vilbois, M. (2015) Exploiting light chains for the scalable generation and platform purification of native human bispecific IgG, Nat Commun. 6, 6113.9. Bacac, M., Colombetti, S., Herter, S., Sam, J., Perro, M., Chen, S., Bianchi, R., Richard, M., Schoenle, A., Nicolini, V., Diggelmann, S., Limani, F., Schlenker, R., Husser, T., Richter, W., Bray-French, K., Hinton, H., Giusti, A. M., Freimoser-Grundschober, A., Lariviere, L., Neumann, C., Klein, C. & Umana, P. (2018) CD20-TCB with Obinutuzumab Pretreatment as Next-Generation Treatment of Hematologic Malignancies, Clin Cancer Res. 24, 4785-4797.10. Strohl, W. R. & Naso, M. (2019) Bispecific T-Cell Redirection versus Chimeric Antigen Receptor (CAR)-T Cells as Approaches to Kill Cancer Cells, Antibodies (Basel). 8.11. Zhukovsky, E. A., Morse, R. J. & Maus, M. V. (2016) Bispecific antibodies and CARs: generalized immunotherapeutics harnessing T cell redirection, Curr Opin Immunol. 40, 24-35.12. Ellerman, D. (2019) Bispecific T-cell engagers: Towards understanding variables influencing the in vitro potency and tumor selectivity and their modulation to enhance their efficacy and safety, Methods. 154, 102-117.13. Labrijn, A. F. (2019) Bispecific Antibodies: New Strategies and Case Studies in Antibody Engineering & Therapeutics 2019 San Diego, CA.14. Finney, H. M. (2019) Bispecific Target Discovery by Screenign a Large "Select, Mix & Assay" Combinatorial Library in Antibody Engineering & Therapeutics 2019 San Diego, CA.15. Joshi, K. (2019) Toward a Deeper Understanding of Bispecific Antibodies in Antibody Engineering & Therapeutics 2019 San Diego, CA.16. Wang, Q., Chen, Y., Park, J., Liu, X., Hu, Y., Wang, T., McFarland, K. & Betenbaugh, M. J. (2019) Design and Production of Bispecific Antibodies, Antibodies (Basel). 8.17. Zhu, Y., Shen, R., Hao, R., Wang, S. & Ho, M. (2020) Highlights of Antibody Engineering & Therapeutics 2019 in San Diego, USA: Bispecific Antibody Design and Clinical Applications, Antibody Therapeutics.18. Labrijn, A. F., Janmaat, M. L., Reichert, J. M. & Parren, P. (2019) Bispecific antibodies: a mechanistic review of the pipeline, Nature reviews Drug discovery. 18, 585-608.19. Bonisch, M., Sellmann, C., Maresch, D., Halbig, C., Becker, S., Toleikis, L., Hock, B. & Ruker, F. (2017) Novel CH1:CL interfaces that enhance correct light chain pairing in heterodimeric bispecific antibodies, Protein Eng Des Sel. 30, 685-696.20. Garfall, A. L. & June, C. H. (2019) Trispecific antibodies offer a third way forward for anticancer immunotherapy, Nature. 575, 450-451.21. Wu, L., Seung, E., Xu, L., Rao, E., Lord, D. M., Wei, R. R., Cortez-Retamozo, V., Ospina, B., Posternak, V., Ulinski, G., Piepenhagen, P., Francesconi, E., El-Murr, N., Beil, C., Kirby, P., Li, A., Fretland, J., Vicente, R., Deng, G., Dabdoubi, T., Cameron, B., Bertrand, T., Ferrari, P., Pouzieux, S., Lemoine, C., Prades, C., Park, A., Qiu, H., Song, Z., Zhang, B., Sun, F., Chiron, M., Rao, S., Radošević, K., Yang, Z.-y. & Nabel, G. J. (2020) Trispecific antibodies enhance the therapeutic efficacy of tumor-directed T cells through T cell receptor co-stimulation, Nature Cancer. 1, 86-98.22. Nabel, G. (2019) Trispecific Antibodies for Treating Cancer and AIDS in Antibody Engineering & Therapeutics 2019 San Diego, CA.23. Chen, D. S. (2019) Defining the Light at the End of the Tunnel: Biologic Problems and Engineering Solutions in Cancer Immunotherapy in Antibody Engineering & Therapeutics 2019 San Diego, CA.24. Budde, E. (2019) Clinical Use of Bispecific Antibodies in Treating B Cell Lymphoma in Antibody Engineering & Therapeutics 2019 San Diego, CA.25. Tesar, M. (2019) CD16A, NK Cell Targeting in Antibody Engineering & Therapeutics 2019 San Diego, CA.26. Haneke, T., Rajkovic, E., Treder, M., Tesar, M. & Koch, J. (2019) Redirected Optimized Cell Killing (ROCK®): A novel multispecific antibody platform for innate immune cell engagement to fight cancer in NK2019 Luxembourg.