预约演示
更新于:2025-09-09

University of California, Davis
更新于:2025-09-09
概览
标签
肿瘤
神经系统疾病
呼吸系统疾病
多肽偶联核素
诊断用放射药物
小分子化药
疾病领域得分
一眼洞穿机构专注的疾病领域

技术平台
公司药物应用最多的技术
靶点
公司最常开发的靶点
关联
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |

靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |


靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |


NCT06774417
Digital Strategies to Advance Help-Seeking in Youth at Clinical High Risk for Developing Psychosis

NCT07076862
Multiparametric Total-Body [18F]F-AraG PET/CT Imaging in Post-Acute Sequelae of SARS-CoV-2 Infection (PASC)
NCT07024498
Coordinating Outpatient bupreNorphiNe for Emergency Care and Continuing Treatment
100 项与 University of California, Davis 相关的临床结果
登录后查看更多信息
登录后查看更多信息
2026-01-01JOURNAL OF AFFECTIVE DISORDERS
Multi-marker cerebral small vessel disease score and risk of incident depression: The Framingham Heart Study
Article
作者: Araujo-Contreras, Robert ; Pinheiro, Adlin Alicia ; Beiser, Alexa ; Himali, Jayandra Jung ; Lioutas, Vasileios ; DeCarli, Charles ; Romero, Jose Rafael ; Seshadri, Sudha ; Sisto, Joseph ; Aparicio, Hugo J ; Ekenze, Oluchi ; Demissie, Serkalem
BACKGROUND:
The vascular depression hypothesis postulates that cerebral small vessel disease (CSVD) contributes to the development of depression. This study examined the relationship between a multi-marker CSVD score derived from brain magnetic resonance imaging (MRI) with incident depression in community-dwelling individuals.
METHODS:
Framingham Heart Study (FHS) participants free of stroke, dementia, and depression with available data on CSVD MRI markers were included. The CSVD score was calculated by summing one point for each individual CSVD marker: covert brain infarcts, cerebral microbleeds, white matter hyperintensities, visible perivascular spaces (PVS) and cortical superficial siderosis, ranging 0-5. Depression was defined as Center for Epidemiological Studies Depression scale score ≥ 16 or antidepressant used. Multivariable logistic regression models were used to relate the CSVD score and individual markers to incident depression. Models were adjusted for age, sex, FHS cohort, time interval between MRI and clinic exam, and regular vascular risk factors.
RESULTS:
Among 1768 participants, (mean age 54.4 ± 12.4 years, 54 % male), 11 % developed depression (median follow-up 6.0 years, IQR: 5.25-6.70). No CSVD score group, or individual marker had a statistically significant association with incident depression. However, both a CSVD score of 3+ (OR: 2.72, 95 %, CI 0.90, 8.19, p = 0.09) and the presence of visible PVS (OR: 1.56, CI 0.97, 2.52, p = 0.07) showed a trend towards increased odds of developing depression.
CONCLUSION:
A high CSVD burden and the presence of visible PVS may be related to incident depression. Validation in larger and more diverse cohorts is required.
2025-12-31DRUG DELIVERY
Safety and biocompatibility of a novel biodegradable aflibercept-drug delivery system in rhesus macaques
Article
作者: Shim, Jaeho ; Mieler, William F. ; Story, Brett D. ; Kang-Mieler, Jennifer J. ; Rudeen, Kayla M. ; Thomasy, Sara M. ; Ferneding, Michelle ; Motta, Monica ; Blandino, Andrew ; Ardon, Monica ; Teixeira, Leandro B. C. ; Roszak, Karolina ; Yiu, Glenn ; Park, Sangwan ; Le, Sophie
A clinical need exists for more effective intravitreal (IVT) drug delivery systems (DDS). This study tested the hypothesis that a novel biodegradable, injectable microsphere-hydrogel drug delivery system loaded with aflibercept (aflibercept-DDS) would exhibit long-term safety and biocompatibility in a non-human primate (NHP) model. We generated aflibercept-loaded poly (lactic-co-glycolic acid) microparticles with a modified double emulsion technique then embedded them into a biodegradable, thermo-responsive poly (ethylene glycol)-co-(L-lactic-acid) diacrylate/N-isopropylacrylamide hydrogel. Aflibercept-DDS (50 µL, 15 µg) was injected into the right eye of 23 healthy rhesus macaques. A complete ophthalmic examination, intraocular pressure (IOP), corneal pachymetry, specular microscopy, A-scan biometry, streak retinoscopy, spectral-domain optical coherence tomography (SD-OCT), fluorescein angiography (FA), and electroretinography (ERG) were performed monthly. Globes from 7 NHPs were histologically examined. Aflibercept-DDS was visualized in the vitreous up to 9 months post-IVT injection, slightly impeding fundoscopy in 4 of 23 eyes; no other consistent abnormalities were appreciated during ophthalmic examination. The IOP and total retinal thickness remained normal in all animals over all timepoints. Central corneal thickness, endothelial cell density, axial globe length, and refractive error did not significantly differ from baseline. Scotopic mixed rod-cone implicit times and amplitudes along with photopic cone response implicit times and amplitudes did not significantly differ from control values. No retinal or choroidal vascular abnormalities were detected with FA and normal retinal architecture was preserved using SD-OCT. Intravitreal injection of a biodegradable aflibercept-DDS was safe and well tolerated in NHPs up to 24 months.
2025-12-31Autophagy reports
Image-based temporal profiling of autophagy-related phenotypes
Article
作者: Thilakaratne, Eshan ; Gangaraju, Ritika ; Adia, Neil Alvin B ; Shah, Priya S ; Beesabathuni, Nitin Sai
Autophagy is a dynamic process critical in maintaining cellular homoeostasis. Dysregulation of autophagy is linked to many diseases and is emerging as a promising therapeutic target. High-throughput methods to characterise autophagy are essential for accelerating drug discovery and characterising mechanisms of action. In this study, we developed a scalable image-based temporal profiling approach to characterise ~900 morphological features at a single cell level with high temporal resolution. We differentiated drug treatments based on morphological profiles using a random forest classifier with ~90% accuracy and identified the key features that govern classification. Additionally, temporal morphological profiles accurately predicted biologically relevant changes in autophagy after perturbation, such as total cargo degraded. Therefore, this study acts as proof-of-principle for using image-based temporal profiling to differentiate autophagy perturbations in a high-throughput manner and has the potential identify biologically relevant autophagy phenotypes. Ultimately, approaches like image-based temporal profiling can accelerate drug discovery.
2025-09-02
·药时空
细胞疗法基因疗法免疫疗法临床申请信使RNA

100 项与 University of California, Davis 相关的药物交易
登录后查看更多信息
100 项与 University of California, Davis 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2025年11月02日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
药物发现
1
9
临床前
临床1期
9
2
临床2期
临床3期
1
14
其他
登录后查看更多信息
当前项目
登录后查看更多信息
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
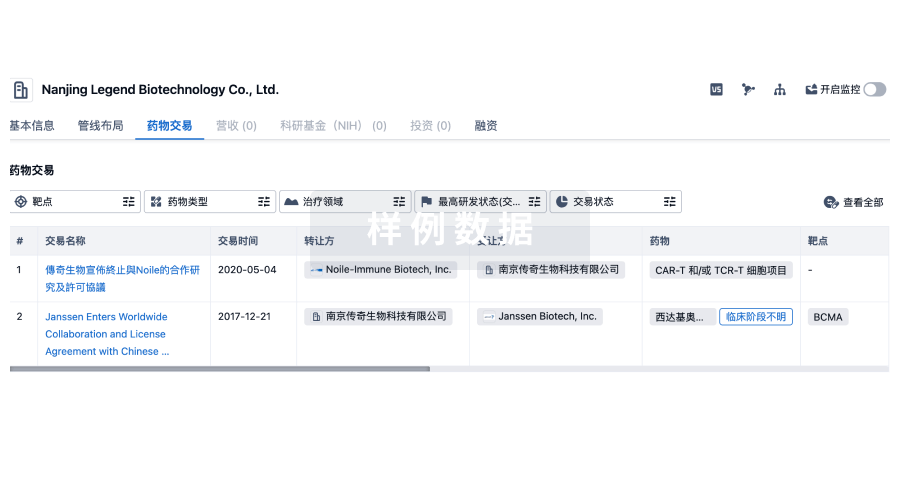
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
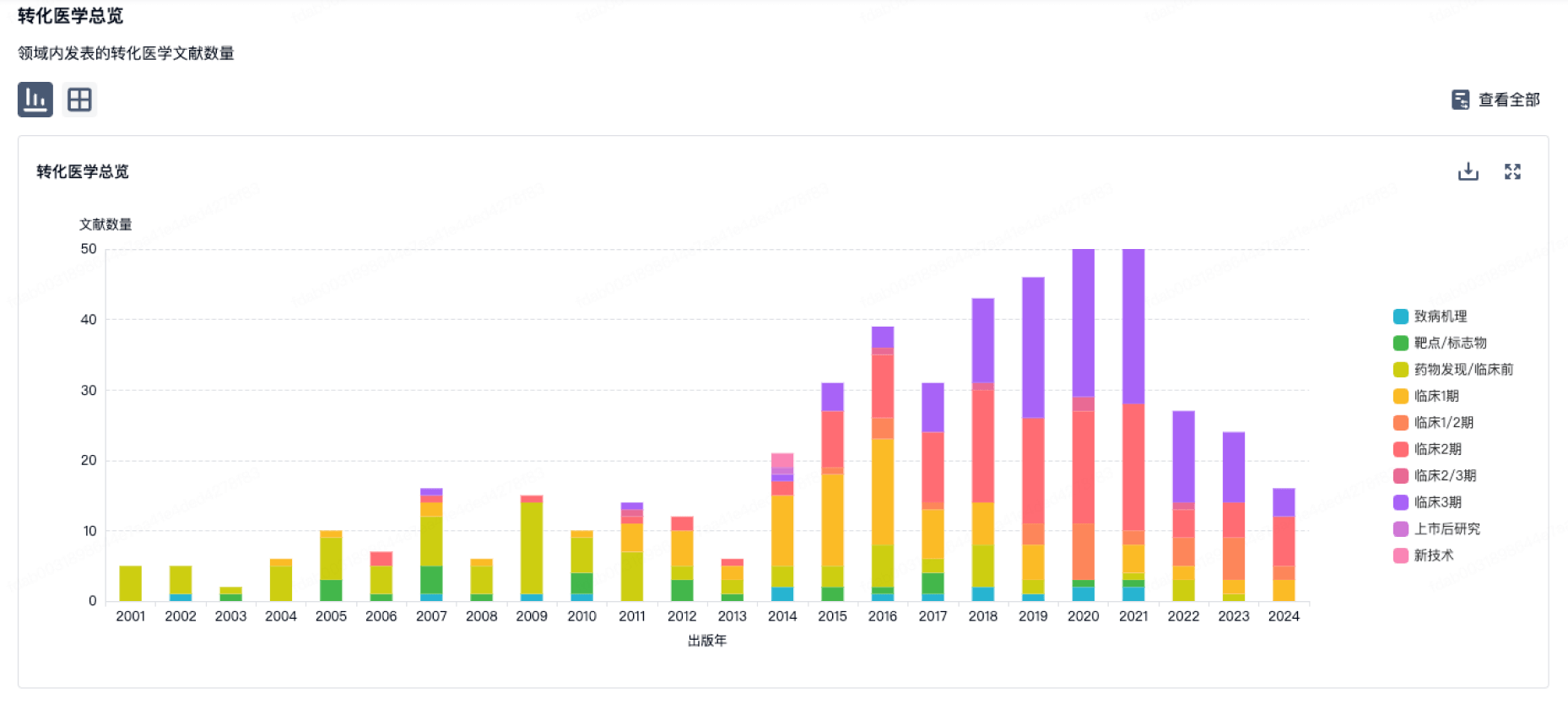
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
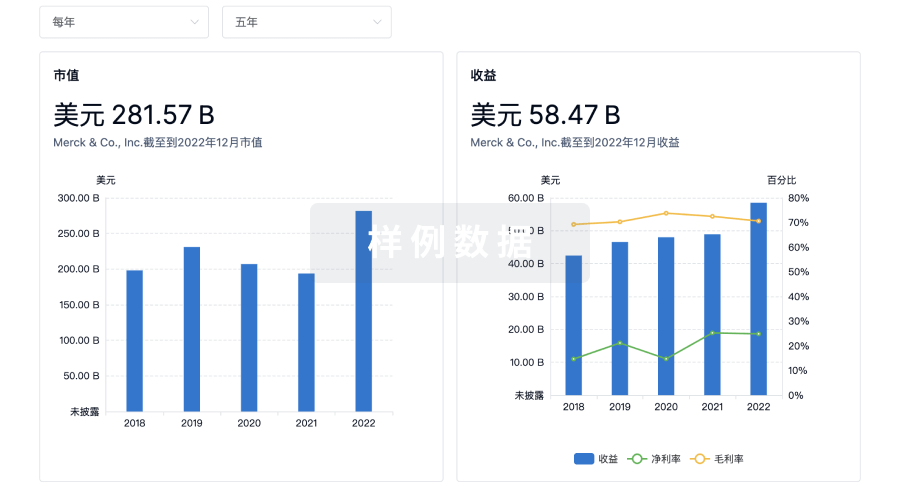
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用