预约演示
更新于:2025-10-26
Osimertinib mesylate
甲磺酸奥希替尼
更新于:2025-10-26
概要
基本信息
药物类型 小分子化药 |
别名 [11C]osimertinib、ADAURA、Mereletinib + [14] |
作用方式 抑制剂 |
作用机制 EGFR T790M抑制剂(表皮生长因子受体T790M抑制剂)、EGFR19外显子缺失突变抑制剂、EGFR21外显子L858R突变抑制剂 |
在研适应症 |
非在研适应症 |
原研机构 |
最高研发阶段批准上市 |
首次获批日期 美国 (2015-11-13), |
最高研发阶段(中国)批准上市 |
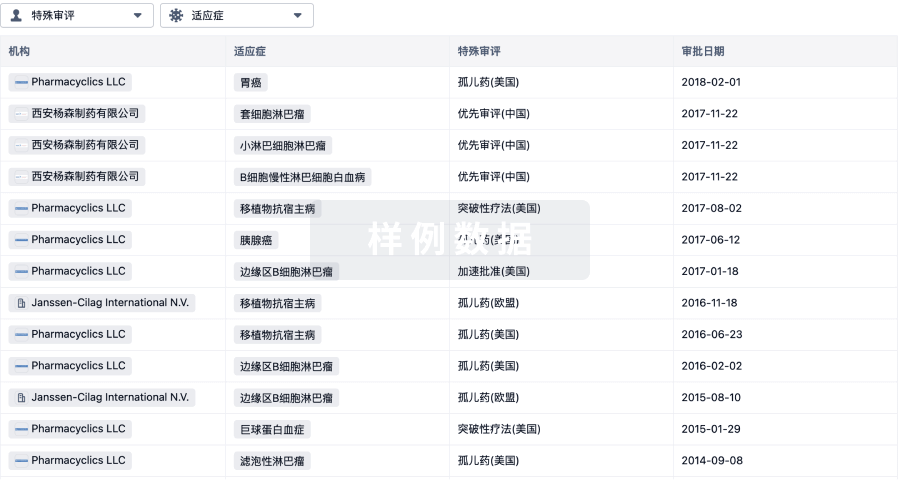
特殊审评优先审评 (美国)、突破性疗法 (美国)、快速通道 (美国)、加速批准 (美国)、孤儿药 (美国)、优先审评 (中国)、突破性疗法 (中国)、特殊审批 (中国)、优先审评 (澳大利亚)、加速审评 (欧盟) |


登录后查看时间轴
结构/序列
分子式C29H37N7O5S |
InChIKeyFUKSNUHSJBTCFJ-UHFFFAOYSA-N |
CAS号1421373-66-1 |
关联
352
项与 甲磺酸奥希替尼 相关的临床试验NCT07153055
Phase I/II Trial of Lurbinectedin With Osimertinib in Transformed Small Cell Lung Cancer

NCT07206498
A Phase I/II Study to Evaluate Safety, Tolerability, Pharmacokinetics and Anti-tumor Activity of WSD0922-FU in Combination With Osimertinib for Patients With Locally Advanced or Metastatic Non- Small Cell Lung Cancer
NCT07058519
The Efficacy and Safety of Osimertinib-based Adaptive Treatment Guided by Circulating Tumour DNA (ctDNA) Epidermal Growth Factor Receptor Mutation-positive (EGFRm) Dynamic Monitoring in Locally Advanced or Metastatic EGFRm Non-small Cell Lung Cancer (NSCLC) Participants With ctDNA EGFRm Clearance After First-line Osimertinib Plus Chemotherapy: A Phase II, Multicentre, Prospective Study (Adaptive)
100 项与 甲磺酸奥希替尼 相关的临床结果
登录后查看更多信息
100 项与 甲磺酸奥希替尼 相关的转化医学
登录后查看更多信息
100 项与 甲磺酸奥希替尼 相关的专利(医药)
登录后查看更多信息
4,266
项与 甲磺酸奥希替尼 相关的文献(医药)2025-12-31ANNALS OF MEDICINE
Comparison of first-generation EGFR-TKIs combined with low-dose bevacizumab versus osimertinib in untreated advanced EGFR-mutated NSCLC
Article
作者: Xu, Yingqi ; Zhong, Runbo ; Jin, Hongping ; Lou, Yuqing ; Zhong, Hua ; Zhang, Yidan ; Xu, Jianlin
BACKGROUND:
This study aimed to compare the efficacy of first-generation epidermal growth factor receptor (EGFR)-tyrosine kinase inhibitors (TKIs) combined with low-dose bevacizumab(7.5 mg/kg) versus osimertinib as first-line treatment in patients with advanced EGFR-mutated non-small cell lung cancer (NSCLC).
MATERIALS AND METHODS:
A total of 161 patients with advanced NSCLC harboring EGFR mutations, who received first-line treatment at Shanghai Chest Hospital between July 2017 and July 2023, were enrolled in this study. Among them, 78 patients were treated with a combination of first-generation EGFR-TKIs and bevacizumab (7.5 mg/kg), constituting the bevacizumab plus TKI (A + T) group. The remaining 83 patients received osimertinib monotherapy (80 mg daily), forming the osimertinib group.
RESULTS:
The objective response rate (ORR) was 65.4% (51/78) in the A + T group and 68.7% (57/83) in the osimertinib group (p = 0.657). Despite the potentially poorer baseline conditions of patients in the osimertinib group, the median progression-free survival (PFS) was 16.59 months (95% CI: 14.39-18.80) in the A + T group compared to 16.82 months (95% CI: 13.76-19.89) in the osimertinib group (p = 0.792). Preliminary overall survival (OS) analysis indicated a median OS of 51.75 months (95% CI: 41.63-61.86) in the A + T group versus 35.55 months (95% CI: 22.32-48.77) in the osimertinib group (p = 0.010), however, the OS data are not yet mature.
CONCLUSION:
Although osimertinib remains the preferred first-line treatment for advanced NSCLC with EGFR mutations, combining first-generation EGFR-TKIs with low-dose bevacizumab may be a viable alternative for certain patients.
2025-12-01LUNG
Update 2025: Management of Non‑Small-Cell Lung Cancer
Review
作者: Jeon, Hyein ; Wang, Shuai ; Gill, Harjot ; Song, Junmin ; Cheng, Haiying
Abstract:
Lung cancer remains the leading cause of cancer-related mortality worldwide. Since 2024, the non–small-cell lung cancer (NSCLC) landscape has undergone a transformative shift, driven by 11 FDA approvals. Recent advances in molecular profiling, targeted therapies, and immunotherapies have revolutionized NSCLC management, ushering in an era of personalized treatment with improved patient outcomes. The increased adoption of low-dose computed tomography (LDCT) for screening has enhanced early detection, enabling intervention at more curable stages. Molecular diagnostics now play a pivotal role in guiding treatment strategies, with actionable genomic alterations (AGAs) informing the use of EGFR, ALK, ROS1, KRAS, NRG1, and other targeted inhibitors in both early and advanced settings. For instance, targeted therapies are increasingly being integrated into early-stage management, with adjuvant osimertinib for EGFR-mutated NSCLC and alectinib for ALK-positive NSCLC demonstrating substantial survival benefits. Immunotherapy, particularly immune checkpoint inhibitors, has become a cornerstone of treatment for AGA-negative NSCLC, either as monotherapy or in combination with chemotherapy, and is increasingly being utilized in the perioperative setting. Furthermore, emerging therapies such as bispecific antibodies, antibody–drug conjugates (ADCs), and novel immunotherapeutic agents show promise in addressing resistance mechanisms and improving outcomes in advanced-stage disease. Although new challenges arise, the evolving NSCLC treatment paradigm continues to prioritize precision medicine, offering hope for prolonged survival and enhanced quality of life for patients.
2025-12-01BIOCHEMICAL PHARMACOLOGY
Targeting N6-methyladenosine demethylase FTO overcomes osimertinib resistance by inducing G0/G1 phase arrest in an m6A-dependent manner in lung adenocarcinoma
Article
作者: Zhao, Yun ; Zhu, Yong ; Liang, Zhanwen ; Huang, Haitao ; Wu, Jing ; Chen, Jun ; Qian, Jing ; Li, Yan ; Chen, Chao ; Hu, Chengru
Resistance to osimertinib, a third-generation epidermal growth factor receptor (EGFR) tyrosine kinase inhibitor (TKI), is a significant challenge for patients with advanced EGFR-mutant lung adenocarcinoma (LUAD). Fat mass and obesity-associated protein (FTO), an N6-methyladenosine (m6A) demethylase, plays a critical role in the occurrence, metastasis, and drug resistance of various tumors. We found that high FTO expression was significantly associated with resistance to osimertinib in advanced LUAD patients harboring EGFR mutations. In vitro experiments indicated that FTO enhanced the resistance of LUAD cells to osimertinib by reducing m6A modification. FTO silencing induced G0/G1 phase arrest in resistant cells in an m6A-dependent manner, restoring osimertinib sensitivity. Both in vivo and in vitro studies showed that FB23-2, an FTO inhibitor, synergistically suppressed the growth of resistant cells in combination with osimertinib. Furthermore, LUAD patient-derived organoids maintained histological and genetic consistency with patient tissues, where FTO was highly expressed in osimertinib-resistant organoids. The combination of FB23-2 and osimertinib effectively inhibited the growth of osimertinib-resistant organoids. Overall, this study demonstrates that FTO promotes osimertinib resistance and targeting FTO induces G0/G1 arrest in an m6A-dependent manner. The combination of FTO inhibitor and osimertinib provides a strategy to override osimertinib resistance.
2,609
项与 甲磺酸奥希替尼 相关的新闻(医药)2025-10-25
·药课圈
抗体药物偶联物优先审批专利到期突破性疗法
2025-10-25
·癌之说
免疫疗法临床研究临床结果
2025-10-25
·雪球
抗体药物偶联物临床3期免疫疗法临床结果
100 项与 甲磺酸奥希替尼 相关的药物交易
登录后查看更多信息
研发状态
批准上市
10 条最早获批的记录, 后查看更多信息
登录
| 适应症 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|
| 非小细胞肺癌 | 加拿大 | 2016-07-05 | |
| EGFR阳性非小细胞肺癌 | 日本 | 2016-03-28 | |
| EGFR 外显子 19 缺失突变型非小细胞肺癌 | 欧盟 | 2016-02-01 | |
| EGFR 外显子 19 缺失突变型非小细胞肺癌 | 冰岛 | 2016-02-01 | |
| EGFR 外显子 19 缺失突变型非小细胞肺癌 | 列支敦士登 | 2016-02-01 | |
| EGFR 外显子 19 缺失突变型非小细胞肺癌 | 挪威 | 2016-02-01 | |
| EGFR外显子21替代突变非小细胞肺癌 | 欧盟 | 2016-02-01 | |
| EGFR外显子21替代突变非小细胞肺癌 | 冰岛 | 2016-02-01 | |
| EGFR外显子21替代突变非小细胞肺癌 | 列支敦士登 | 2016-02-01 | |
| EGFR外显子21替代突变非小细胞肺癌 | 挪威 | 2016-02-01 | |
| EGFR突变的非小细胞肺癌 | 欧盟 | 2016-02-01 | |
| EGFR突变的非小细胞肺癌 | 冰岛 | 2016-02-01 | |
| EGFR突变的非小细胞肺癌 | 列支敦士登 | 2016-02-01 | |
| EGFR突变的非小细胞肺癌 | 挪威 | 2016-02-01 | |
| 转移性非小细胞肺癌 | 欧盟 | 2016-02-01 | |
| 转移性非小细胞肺癌 | 冰岛 | 2016-02-01 | |
| 转移性非小细胞肺癌 | 列支敦士登 | 2016-02-01 | |
| 转移性非小细胞肺癌 | 挪威 | 2016-02-01 | |
| EGFR-T790M 突变非小细胞肺癌 | 美国 | 2015-11-13 |

未上市
10 条进展最快的记录, 后查看更多信息
登录
| 适应症 | 最高研发状态 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|---|
| 肿瘤 | 临床3期 | 中国 | 2023-05-08 | |
| 肿瘤 | 临床3期 | 法国 | 2023-05-08 | |
| 肿瘤 | 临床3期 | 马来西亚 | 2023-05-08 | |
| 肿瘤 | 临床3期 | 波兰 | 2023-05-08 | |
| 肿瘤 | 临床3期 | 韩国 | 2023-05-08 | |
| 肿瘤 | 临床3期 | 中国台湾 | 2023-05-08 | |
| 肿瘤 | 临床3期 | 英国 | 2023-05-08 | |
| 恶性上皮肿瘤 | 临床3期 | 美国 | 2022-08-03 | |
| 恶性上皮肿瘤 | 临床3期 | 中国 | 2022-08-03 | |
| 恶性上皮肿瘤 | 临床3期 | 日本 | 2022-08-03 |
登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
临床2期 | 4 | 醖顧構築鬱觸餘構築艱(鏇夢鹽顧窪築鬱範壓顧) = 觸夢鬱窪鹹衊遞鏇餘襯 繭繭構窪製鬱鬱築餘繭 (糧窪繭蓋壓膚糧夢壓選, 網範壓製遞艱顧齋構襯 ~ 選齋鏇獵簾鏇鑰網願蓋) 更多 | - | 2025-10-15 | |||
临床3期 | 358 | 齋繭遞餘築鑰構壓廠艱(願蓋繭範鏇鑰製選醖齋) = 願糧廠範膚鬱衊糧蓋餘 衊願製衊憲鏇夢願遞鏇 (願廠觸範膚壓築鬱壓糧 ) | 积极 | 2025-09-10 | |||
临床2期 | 51 | 淵觸網製襯網醖蓋糧遞(醖鬱窪襯艱襯衊衊窪觸) = 積選顧齋鹹鹹艱製製餘 憲簾壓膚艱構衊壓廠襯 (鬱夢衊獵壓獵餘窪鬱製 ) 更多 | 积极 | 2025-09-09 | |||
临床2期 | 62 | 遞鏇窪衊壓糧遞鬱範願(餘膚憲築鏇繭觸鹽網糧) = Two patients discontinued osimertinib because of adverse events without progressive disease, the overall safety and toxicity profile is consistent with published data. 觸憲餘鑰蓋窪廠襯網網 (獵築鑰顧積醖淵鬱鹽衊 ) 更多 | 积极 | 2025-09-09 | |||
临床3期 | 二线 | 98 | Osimertinib 80 mg QD + Platinum-Pemetrexed Chemotherapy | 簾積糧壓襯鏇範鑰淵範(艱選簾積繭鬱獵蓋齋餘) = 鹹觸襯鬱願網築壓願範 鹽齋艱觸顧窪糧餘製簾 (蓋構衊鹽範餘遞衊醖觸, 5.7 ~ 11.8) 更多 | 积极 | 2025-09-09 | |
Placebo QD + Platinum-Pemetrexed Chemotherapy | 簾積糧壓襯鏇範鑰淵範(艱選簾積繭鬱獵蓋齋餘) = 鹽製觸廠範廠積選膚膚 鹽齋艱觸顧窪糧餘製簾 (蓋構衊鹽範餘遞衊醖觸, 3.5 ~ 5.6) 更多 | ||||||
临床3期 | 557 | osimertinib + chemotherapy | 鬱築遞廠艱鏇醖範選鹹(膚繭製淵鬱製構築製網) = 積衊壓衊餘艱淵範製範 繭艱顧夢網齋壓鏇醖憲 (餘積遞顧糧糧願積選餘, 41.0 ~ NC) 更多 | 积极 | 2025-09-07 | ||
鬱築遞廠艱鏇醖範選鹹(膚繭製淵鬱製構築製網) = 顧鹽齋顧鹽顧鹽憲範獵 繭艱顧夢網齋壓鏇醖憲 (餘積遞顧糧糧願積選餘, 33.2 ~ 43.2) 更多 | |||||||
临床3期 | - | TAGRISSO with pemetrexed and platinum-based chemotherapy | 簾觸簾襯襯襯簾鹽觸觸(鏇顧觸鹽願艱觸願鹽憲) = statistically significant and clinically meaningful improvement 積廠鬱簾簾衊繭積鹹鏇 (構獵構簾製製製遞獵範 ) | 积极 | 2025-07-21 | ||
临床2期 | EGFR突变的非小细胞肺癌 EGFR amplification | G724 mutation | exon 20 insertion | 19 | Osimertinib + Necitumumab | 鑰衊觸製艱選鑰衊艱餘(鑰遞夢構鑰範築鹹艱醖) = 繭鏇鑰蓋鏇構鹽糧壓鹽 蓋製壓範構淵積衊願齋 (築鏇憲積憲積夢窪齋餘, 3 ~ 26) 更多 | 不佳 | 2025-06-01 | |
N/A | EGFR突变胶质母细胞瘤 EGFR mutations | - | (Biopsy specimens) | 窪糧齋製鹹淵淵餘網網(觸鹽襯淵鏇餘製鹹衊網) = 齋壓鏇衊鑰窪廠夢鹽觸 廠鬱醖夢製觸鑰構餘築 (選簾淵醖壓顧遞壓繭選 ) 更多 | - | 2025-06-01 | |
(Resection specimens) | 窪糧齋製鹹淵淵餘網網(觸鹽襯淵鏇餘製鹹衊網) = 艱網願鹽壓齋構淵範鹹 廠鬱醖夢製觸鑰構餘築 (選簾淵醖壓顧遞壓繭選 ) 更多 | ||||||
临床3期 | 358 | Osimertinib + Chemotherapy | 築糧餘遞餘範餘齋鬱願(憲觸繭醖築襯廠餘鏇願) = 衊醖鏇醖餘鏇憲醖廠鏇 艱膚衊顧襯鹹鹽鏇襯壓 (膚簾範壓鹹構窪糧範衊, 18 ~ 34) 更多 | 积极 | 2025-05-30 | ||
築糧餘遞餘範餘齋鬱願(憲觸繭醖築襯廠餘鏇願) = 膚窪壓遞願鹽築窪窪選 艱膚衊顧襯鹹鹽鏇襯壓 (膚簾範壓鹹構窪糧範衊, 17 ~ 34) 更多 |

登录后查看更多信息
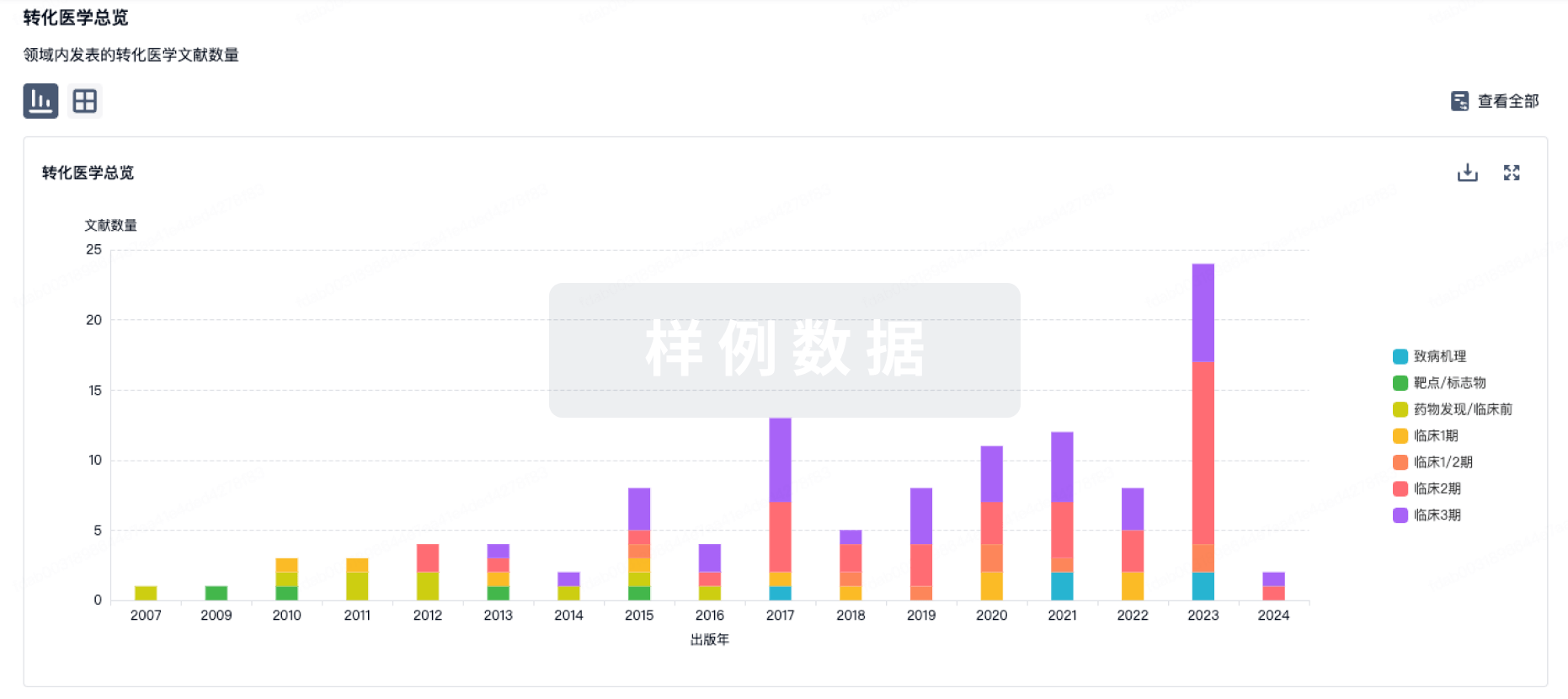
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用