预约演示
更新于:2025-06-24

Rui Therapeutics Co., Ltd
深圳恩瑞恺诺生物技术有限公司|控股公司|2021|中国广东省
深圳恩瑞恺诺生物技术有限公司|控股公司|2021|中国广东省
更新于:2025-06-24
概览
标签
免疫系统疾病
皮肤和肌肉骨骼疾病
血液及淋巴系统疾病
CAR-NK
CAR-T
疾病领域得分
一眼洞穿机构专注的疾病领域

暂无数据
技术平台
公司药物应用最多的技术
暂无数据
靶点
公司最常开发的靶点
暂无数据
| 排名前五的药物类型 | 数量 |
|---|---|
| CAR-NK | 8 |
| CAR-T | 1 |
| 排名前五的靶点 | 数量 |
|---|---|
| BCMA x CD19 | 1 |
| CD19(B淋巴细胞抗原CD19) | 1 |
关联
9
项与 深圳恩瑞恺诺生物技术有限公司 相关的药物靶点 |
作用机制 CD19抑制剂 [+2] |
在研机构 |
原研机构 |
非在研适应症- |
最高研发阶段临床1期 |
首次获批国家/地区- |
首次获批日期- |
作用机制 BCMA调节剂 [+1] |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症- |
最高研发阶段早期临床1期 |
首次获批国家/地区- |
首次获批日期- |
靶点- |
作用机制- |
在研机构 |
原研机构 |
在研适应症- |
非在研适应症- |
最高研发阶段临床阶段不明 |
首次获批国家/地区- |
首次获批日期- |
7
项与 深圳恩瑞恺诺生物技术有限公司 相关的临床试验NCT06976437
An Exploratory Clinical Study on the Safety and Efficacy of Allogeneic CD19/BCMA CAR-T Cell Treatment for Relapsed/ Refractory B-cell or Plasma Cell-derived Malignant Tumors
A single arm, open-label pilot study is designed to determine the safety and efficacy of CD19 and B-cell maturation antigen (BCMA) targeted allogenic CAR-T cells (RN1101) in patients with relapsed/refractory B-cell or plasma cell-derived malignant tumors. 21 patients are planned to be enrolled in the dose-escalation trial. The primary objective of the study is to evaluation of the safety and feasibility of RN1101 for the treatment of relapsed/refractory B-cell or plasma cell-derived malignant tumors. The secondary objective is to evaluate the efficacy of RN1101 for the treatment of relapsed/refractory B-cell or plasma cell-derived malignant tumors. The exploratory objective is to evaluate expansion, persistence and ability of RN1101 to deplete CD19 or BCMA positive cells in patients with relapsed/refractory B-cell or plasma cell-derived malignant tumors.
开始日期2025-05-28 |
申办/合作机构 [+1] |
NCT06613490
An Exploratory Clinical Study of the Safety and Efficacy of CD19 Chimeric Antigen Receptor NK Cells for the Treatment of Refractory Antisynthetase Antibody Syndrome and Rheumatoid Arthritis
A single-center, open-label dose-escalation design to evaluate the safety and efficacy of 3 infusions of anti CD19 CAR NK cells (KN5501), as well as the expansion and persistence of KN5501 in patients with refractory antisynthetase antibody syndrome (ASyS) and rheumatoid arthritis (RA); To evaluate the ability of KN5501 to clear CD19-positive B cells in patients to determine the feasibility of KN5501 for the treatment of refractory ASyS and or RA.
开始日期2025-05-20 |
申办/合作机构 [+1] |
NCT06337474
An Exploratory Clinical Study of the Safety and Efficacy of CD19 Chimeric Antigen Receptor NK Cell Injections for the Treatment of Refractory Primary Immune Thrombocytopenia
A single arm, open-label pilot study is designed to determine the safety and effectiveness of CD19 CAR NK cells (KN5501) in patients with refractory immune thrombocytopenia. 9 patients are planned to be enrolled in the dose-escalation trial (9×10^9 cells, 13.5×10^9 cells). The primary objective of the study is to evaluation of the safety and feasibility of KN5501 for the treatment of relapsed/refractory B-cell related autoimmune diseases. The secondary objective is to evaluate evaluation of KN5501 for the treatment of refractory immune thrombocytopenia. The exploratory objective is to evaluate expansion, persistence and ability to deplete CD19 positive B cells of KN5501 in patients with refractory immune thrombocytopenia.
开始日期2024-08-30 |
申办/合作机构 常熟市第二人民医院 [+1] |
100 项与 深圳恩瑞恺诺生物技术有限公司 相关的临床结果
登录后查看更多信息
0 项与 深圳恩瑞恺诺生物技术有限公司 相关的专利(医药)
登录后查看更多信息
20
项与 深圳恩瑞恺诺生物技术有限公司 相关的新闻(医药)2025-06-22
·信狐药迅
本周药品注册受理数据,分门别类呈现,一目了然。(6.16-6.22)新药上市申请无新药临床申请药品名称企业注册分类受理号CX1440胶囊杭州邦顺制药有限公司1CXHL2500603CX1440胶囊杭州邦顺制药有限公司1CXHL2500602HSK47388片海思科医药集团股份有限公司1CXHL2500601HSK47388片海思科医药集团股份有限公司1CXHL2500600磷酸泊沃昔替尼片海南德镁医药科技有限责任公司1CXHL2500599磷酸泊沃昔替尼片海南德镁医药科技有限责任公司1CXHL2500598磷酸泊沃昔替尼片海南德镁医药科技有限责任公司1CXHL2500597BAY 3713372 片拜耳医药保健有限公司1CXHL2500596BAY 3713372 片拜耳医药保健有限公司1CXHL2500595YH004胶囊杭州禹泓医药科技有限公司1CXHL2500592丁二酸复瑞替尼胶囊复星万邦(江苏)医药集团有限公司1CXHL2500591F-02-2-Na片广东横琴新创益生物医药有限公司1CXHL2500589F-02-2-Na片广东横琴新创益生物医药有限公司1CXHL2500588F-02-2-Na片湘北威尔曼制药股份有限公司1CXHL2500587SIM0508片上海先祥医药科技有限公司1CXHL2500584SIM0508片上海先祥医药科技有限公司1CXHL2500583ABSK043片上海和誉生物医药科技有限公司1CXHL2500582ABSK043片上海和誉生物医药科技有限公司1CXHL2500581雷美替胺双释片南京三迭纪医药科技有限公司2.2CXHL2500590FCZR博济医药科技股份有限公司2.2;2.4CXHL2500594FCZR博济医药科技股份有限公司2.2;2.4CXHL2500593注射用西罗莫司(白蛋白结合型)石药集团中奇制药技术(石家庄)有限公司2.2;2.4CXHL2500586奥曲肽长效注射液石药集团中奇制药技术(石家庄)有限公司2.2;2.4CXHL2500585重组带状疱疹疫苗(CHO细胞) (复合佐剂)江苏立维斯德生物技术有限公司1.3CXSL2500498T3011疱疹病毒注射液苏州亦诺微医药科技有限公司1CXSL2500502多抗原自体免疫细胞注射液恒瑞源正(上海)生物科技有限公司1CXSL2500501CM336注射液康诺亚生物医药科技(成都)有限公司1CXSL2500503HX001注射液武汉厚先生物医药有限公司1CXSL2500499SA102-CAR-T注射液沈阳三生制药有限责任公司1CXSL2500497JLM019注射液杰库(上海)生物医药研究有限公司1CXSL2500496KN5501细胞注射液深圳恩瑞恺诺生物技术有限公司1CXSL2500488人脐带间充质干细胞注射液北京再生生物科技研究院有限公司1CXSL2500494脐带间充质干细胞注射液中科聚研(北京)生物科技有限公司1CXSL2500492注射用HS-20093上海翰森生物医药科技有限公司1CXSL2500490SY-005注射液苏州亚宝药物研发有限公司1CXSL2500489SHR-8068注射液苏州盛迪亚生物医药有限公司2.2CXSL2500495怡培生长激素注射液厦门特宝生物工程股份有限公司2.2CXSL2500493阿得贝利单抗注射液上海盛迪医药有限公司2.2CXSL2500491仿制药申请药品名称企业注册分类受理号小儿多种维生素注射液(13)亚宝药业集团股份有限公司3CYHS2502241比拉斯汀口崩片四川科伦药业股份有限公司3CYHS2502229丙酸氟替卡松吸入粉雾剂润生药业有限公司3CYHS2502219苯溴马隆片浙江华海制药科技有限公司3CYHS2502226氨基酸(15)腹膜透析液石家庄四药有限公司3CYHS2502220二硫化硒洗剂福元药业有限公司3CYHS2502216硝普钠注射液海南倍特药业有限公司3CYHS2502213硝普钠注射液广州世济医药科技有限公司3CYHS2502228膦甲酸钠注射液海南如康生物科技有限公司3CYHS2502204盐酸曲唑酮片合肥英太制药有限公司3CYHS2502177盐酸曲唑酮片合肥英太制药有限公司3CYHS2502176盐酸纳美芬注射液石家庄四药有限公司3CYHS2502174比拉斯汀口服溶液河南君善生物技术有限公司3CYHS2502169托吡酯口服溶液广州汉光药业股份有限公司3CYHS2502194硫代硫酸钠注射液中润药业有限公司3CYHS2502193硫代硫酸钠注射液中润药业有限公司3CYHS2502192盐酸左沙丁胺醇雾化吸入溶液广州南新制药有限公司3CYHS2502188帕拉米韦注射液海南惠普森医药生物技术有限公司3CYHS2502187聚乙二醇4000散海南允鼎医药科技有限公司3CYHS2502186艾曲泊帕乙醇胺干混悬剂山东则正医药技术有限公司3CYHS2502182硝普钠注射液仁合益康汇泽药业河北有限公司3CYHS2502175氨磺必利口服溶液西洲医药科技(浙江)有限公司3CYHS2502146琥珀酸亚铁片郑州泰丰制药有限公司3CYHS2502145西格列汀二甲双胍缓释片保定天浩制药有限公司3CYHS2502158小儿复方氨基酸注射液(19AA-I)燃点(南京)生物医药科技有限公司3CYHS2502157加巴喷丁口服溶液海南斯达制药有限公司3CYHS2502156乙酰半胱氨酸片华益药业科技(安徽)有限公司3CYHS2502155盐酸精氨酸注射液石家庄四药有限公司3CYHS2502154草酸艾司西酞普兰口服溶液武汉益博元医药科技有限公司3CYHS2502162硫酸长春新碱注射液海南倍特药业有限公司3CYHS2502160二十碳五烯酸乙酯软胶囊浙江沣华医药科技有限公司4CYHS2502243乙酰半胱氨酸注射液江西银涛药业股份有限公司4CYHS2502242苯磺酸左氨氯地平片苏州俞氏药业有限公司4CYHS2502240苯磺酸左氨氯地平片苏州俞氏药业有限公司4CYHS2502239注射用甲苯磺酸奥马环素苏州天马医药集团天吉生物制药有限公司4CYHS2502238氧(气态)渭南市临渭区新永安医用氧有限责任公司4CYHS2502237甲钴胺片山西汾河制药有限公司4CYHS2502236枸橼酸钠血滤置换液广东科泓药业有限公司4CYHS2502235复方氨基酸注射液(18AA-VII)辰欣药业股份有限公司4CYHS2502233乙酰半胱氨酸注射液安徽金太阳生化药业有限公司4CYHS2502232注射用甲苯磺酸奥马环素海南合瑞制药股份有限公司4CYHS2502231硫酸艾沙康唑胶囊福安药业集团庆余堂制药有限公司4CYHS2502230氨吡啶缓释片石家庄四药有限公司4CYHS2502225溴芬酸钠滴眼液盈科瑞(珠海金湾)制药有限公司4CYHS2502224艾拉莫德片河北龙海药业有限公司4CYHS2502223硫酸氨基葡萄糖胶囊白云山东泰商丘药业有限公司4CYHS2502222瑞维那新吸入溶液广州大光制药有限公司4CYHS2502221聚乙烯醇滴眼液瑞海医药科技(山东)有限公司4CYHS2502218二十碳五烯酸乙酯软胶囊海南斯达制药有限公司4CYHS2502217二十碳五烯酸乙酯软胶囊苏州中化药品工业有限公司4CYHS2502215二十碳五烯酸乙酯软胶囊苏州中化药品工业有限公司4CYHS2502214聚乙烯醇滴眼液广州仁恒医药科技股份有限公司4CYHS2502212乙酰半胱氨酸注射液峨眉山通惠制药有限公司4CYHS2502211蛋白琥珀酸铁口服溶液山东新时代药业有限公司4CYHS2502210乙酰半胱氨酸注射液江苏宝东医药科技有限公司4CYHS2502209硫酸氨基葡萄糖胶囊安徽泰恩康制药有限公司4CYHS2502208蛋白琥珀酸铁口服溶液青岛黄海制药有限责任公司4CYHS2502227瑞维那新吸入溶液齐鲁制药有限公司4CYHS2502234达格列净二甲双胍缓释片(III)广州大光制药有限公司4CYHS2502207达格列净二甲双胍缓释片(II)广州大光制药有限公司4CYHS2502206达格列净二甲双胍缓释片(I)广州大光制药有限公司4CYHS2502205乌帕替尼缓释片重庆圣华曦药业股份有限公司4CYHS2502203他达拉非片海南赛立克药业有限公司4CYHS2502202他达拉非片海南赛立克药业有限公司4CYHS2502201他达拉非片海南赛立克药业有限公司4CYHS2502200盐酸戊乙奎醚注射液湖南一格制药有限公司4CYHS2502198盐酸戊乙奎醚注射液湖南一格制药有限公司4CYHS2502197贝美前列素滴眼液北京北陆药业股份有限公司4CYHS2502196乙酰半胱氨酸注射液安徽茂康药业有限公司4CYHS2502199瑞维那新吸入溶液四川普锐特药业有限公司4CYHS2502195盐酸曲唑酮缓释片华益药业科技(安徽)有限公司4CYHS2502179盐酸曲唑酮缓释片华益药业科技(安徽)有限公司4CYHS2502178中/长链脂肪乳注射液(C8-24Ve)江苏明德制药有限公司4CYHS2502173甲硝唑凝胶杭州和泽坤元药业有限公司4CYHS2502172盐酸乙哌立松片康普药业股份有限公司4CYHS2502170阿卡波糖片北京双鹭药业股份有限公司4CYHS2502168阿卡波糖片北京双鹭药业股份有限公司4CYHS2502167依折麦布辛伐他汀片安徽佳和药业有限公司4CYHS2502166草酸艾司西酞普兰片湖南千金湘江药业股份有限公司4CYHS2502191草酸艾司西酞普兰片湖南千金湘江药业股份有限公司4CYHS2502190草酸艾司西酞普兰片湖南千金湘江药业股份有限公司4CYHS2502189苯磺顺阿曲库铵注射液大道隆达(北京)医药科技发展有限公司4CYHS2502184雌二醇地屈孕酮片扬州奥锐特药业有限公司4CYHS2502183奥美沙坦酯氨氯地平片裕松源药业有限公司4CYHS2502171乙酰半胱氨酸注射液福安药业集团庆余堂制药有限公司4CYHS2502185盐酸氨溴索口服溶液石家庄凯达生物工程有限公司4CYHS2502181盐酸氨溴索口服溶液石家庄凯达生物工程有限公司4CYHS2502180恩格列净片浙江江北药业有限公司4CYHS2502147普瑞巴林胶囊石家庄市华新药业有限责任公司4CYHS2502144普瑞巴林胶囊石家庄市华新药业有限责任公司4CYHS2502143吸入用乙酰半胱氨酸溶液合肥力成药业有限公司4CYHS2502159注射用甲苯磺酸奥马环素四川美大康华康药业有限公司4CYHS2502153培哚普利叔丁胺片华益药业科技(安徽)有限公司4CYHS2502152培哚普利叔丁胺片华益药业科技(安徽)有限公司4CYHS2502151磷酸奥司他韦胶囊江苏诚康药业有限公司4CYHS2502150甲磺酸酚妥拉明注射液海南倍康医药科技有限公司4CYHS2502149氧(液态)石家庄陕鼓气体有限公司4CYHS2502148米格列醇片武汉维奥制药有限公司4CYHS2502165依折麦布片湖南千金湘江药业股份有限公司4CYHS2502164白消安注射液海南倍特药业有限公司4CYHS2502161溴芬酸钠滴眼液山东海山药业有限公司4CYHS2502163达格列净二甲双胍缓释片(I)天津力生制药股份有限公司4CYHS2502142雷珠单抗注射液杰科(天津)生物医药有限公司3.3CXSS2500067德谷门冬双胰岛素注射液宜昌东阳光长江药业股份有限公司3.3CXSS2500066德谷门冬双胰岛素注射液宜昌东阳光长江药业股份有限公司3.3CXSS2500065噁拉戈利片金陵药业股份有限公司南京金陵制药厂3CYHL2500118噁拉戈利片金陵药业股份有限公司南京金陵制药厂3CYHL2500117左甲状腺素钠口服溶液江苏贝美制药有限公司3CYHL2500116甲磺酸酚妥拉明滴眼液广州大光制药有限公司3CYHL2500114黄体酮阴道缓释凝胶河北新张药股份有限公司4CYHL2500115SYS6036注射液石药集团巨石生物制药有限公司3.3CXSL2500500进口申请药品名称企业注册分类受理号紫杉醇口服溶液DAEHWA PHARMACEUTICAL CO.,LTD.2.4JXHS2500073紫杉醇口服溶液DAEHWA PHARMACEUTICAL CO.,LTD.2.4JXHS2500072紫杉醇口服溶液DAEHWA PHARMACEUTICAL CO.,LTD.2.4JXHS2500071马昔腾坦分散片Janssen-Cilag International NV5.1JXHS2500070匹妥布替尼片Eli Lilly Nederland B.V.5.1JXHS2500069匹妥布替尼片Eli Lilly Nederland B.V.5.1JXHS2500068Centanafadine持释胶囊Otsuka Pharmaceutical Co.,Ltd.1JXHL2500152Centanafadine持释胶囊Otsuka Pharmaceutical Co.,Ltd.1JXHL2500151Centanafadine持释胶囊Otsuka Pharmaceutical Co.,Ltd.1JXHL2500150PF-07248144片Pfizer Inc.1JXHL2500149PF-07248144片Pfizer Inc.1JXHL2500148BI 907828片Boehringer Ingelheim International GmbH1JXHL2500147BI 907828片Boehringer Ingelheim International GmbH1JXHL2500146BI 907828片Boehringer Ingelheim International GmbH1JXHL2500145AMG 133Amgen Inc.1JXSL2500109AMG 133Amgen Inc.1JXSL2500108AMG 133Amgen Inc.1JXSL2500107AMG 133Amgen Inc.1JXSL2500106AMG 133Amgen Inc.1JXSL2500105AMG 133Amgen Inc.1JXSL2500104AZD8205AstraZeneca AB1JXSL2500103AZD6621AstraZeneca AB1JXSL2500090AMG 133Amgen Inc.1JXSL2500096AMG 133Amgen Inc.1JXSL2500095AMG 133Amgen Inc.1JXSL2500094AMG 133Amgen Inc.1JXSL2500093AMG 133Amgen Inc.1JXSL2500092AMG 133Amgen Inc.1JXSL2500091注射用贝林妥欧单抗(皮下注射)Amgen Inc.2.1JXSL2500098注射用贝林妥欧单抗(皮下注射)Amgen Inc.2.1JXSL2500097注射用德曲妥珠单抗Daiichi Sankyo, Inc.2.2JXSL2500101比奇珠单抗注射液UCB Pharma S.A.2.2JXSL2500100比奇珠单抗注射液UCB Pharma S.A.2.2JXSL2500099DS-1062aDaiichi Sankyo, Inc.2.2JXSL2500102中药相关申请药品名称企业注册分类受理号通窍静眩颗粒九华华源药业股份有限公司1.1CXZL2500040注:绿色字体部分为潜在首仿品种;不包含原料药、医用氧、注射用水、氯化钠或葡萄糖注射液等申请,不包含再注册、一次性进口、技术转移、复审申请。
疫苗申请上市
2024-11-27
·药智网
10月25日,上海一女子因患系统性红斑狼疮,后因各种原因导致疾病控制不佳,最终导致肾衰竭,需要常年透析维持生命,最终赴瑞士接受安乐死。
10月29日,葛兰素史克(GSK)与恩沐生物达成最新协议,将以高达8.5亿美元的总金额,从恩沐生物收购一种名为CMG1A46的新药,而GSK表示这款新药或将成为破解红斑狼疮这一顽疾的关键。
11月5日,邦耀生物宣布,其开发的“靶向CD19非病毒PD1定点整合CAR-T细胞注射液”的临床试验申请(IND)获得中国国家药品监督管理局药品审评中心(CDE)批准,适应证恰好就是“中度或重度难治性系统性红斑狼疮(SLE)”
短短半月,一边是患者选择毅然赴死,一边是药企加速推进新药上市,系统性红斑狼疮(简称SLE)领域为何会出现如此戏剧性的一幕?
更关键的是,现阶段的SLE治疗究竟是怎样的格局,临床上迫切的临床需求是什么?哪些突破性创新疗法将在未来阻止更多悲剧的发生?
不死的癌症
SLE并非无药可救
所谓系统性红斑狼疮,其发病机制复杂、临床表现多样,目前其发病机制尚未完全阐明,是自免系统疾病中较突出的疾病类型,突出表现为自身免疫性炎症,血清中出现以抗核抗体(ANA)为代表的多种自身抗体及多器官和系统受累,一般情况下,SLE好发于育龄期女性,亚洲地区患病率约为2.5~9.9/10万。
治疗方面,虽然目前仍没有明确有效的根治方法,临床上仍以控制SLE疾病活动,改善临床症状,缓解疾病或达到最低疾病活动度为主要目的,但随着科学家对SLE的研究深入,其治疗方式也从最初的部分缓解,逐渐实现了如今的完全缓解。
图片来源:医学界风湿免疫频道
而根据药物作用机制,SLE的药物治疗则又可以分为抗炎治疗、免疫抑制治疗与靶向治疗三个主要方向:
抗炎治疗:通常情况下,作为贯穿了SLE整个病程的基本用药,代表药物有非甾体抗炎药、羟氯喹与糖皮质激素,其中羟氯喹之类的抗疟药,作为核心用药原则通常建议患者优先使用,而当病情控制不佳后再逐渐加入激素治疗,其中糖皮质激素作为临床一线用药,能有效抑制结缔组织水平的提升,并且能够调控组胺物质水平,是现阶段SLE治疗中最重要的药物。
免疫抑制治疗:其主要是通过抑制机体异常免疫表达来控制体内细胞及体液免疫应答,减轻组织免疫损伤,机制上主要包括清除致敏淋巴细胞、清除机体异常免疫反应中的淋巴细胞、清除非特异炎症中的非淋巴样细胞和抑制残存的淋巴样细胞的功能,代表药物有环磷酰胺、硫唑嘌呤、甲氨蝶呤等。
生物靶向制剂:作为近年来迅速发展的治疗方式,其核心机制主要包括抑制B和T淋巴细胞的活化,阻断免疫细胞间的相互作用,以及抑制细胞因子的产生等,相较其他疗法,其特点在于可作用于特定的病变部位,通过增加目标局部的药物浓度来减轻药物对正常组织和细胞的毒性作用,也提高了治疗的特异性。
原则上来讲,SLE无论过去还是未来,其都并非一款绝对意义上的绝症,只是其特殊的致病机制所导致的治疗困难,且因为现阶段治疗药物普遍存在的较强的副作用,严重影响了患者的生存质量,因此目前SLE治疗上正在逐渐由抗炎与免疫抑制剂向靶向抑制剂演变。
因此,以“激素替代”为方向新药研发也在稳步进行中,成为众多药企争相角逐的战场。
研发火热
SLE在研新药活跃
据研究资料显示,近年来随着越来越多SLE发病机制的关键通路被揭示,如T、B淋巴细胞受体信号通路、细胞因子JAK-STAT信号通路、Toll样受体信号通路等,显著推动了多种靶向治疗如生物制剂、小分子制剂、miRNA类似物以及基因工程T细胞免疫疗法在SLE中的开发及临床应用,目前全球范围内已有多款治疗SLE药物出现。
据不完全统计,截至目前,全球共有279条在研管线用于治疗系统性红斑狼疮适应症,其中活跃管线(近3年有研究进展)共有180余条,占比62%,高于创新药领域平均活跃占比,充分显示了全球新药对SLE领域的研发热情。
数据来源:药智数据、公开数据整理
而从在研SLE新药的临床阶段来看,处于临床II期的管线数量最多,共计88款(活跃管线61款);其次是临床I期,共有80款(活跃管线56款);而临床III期,全球虽仅有21款药物进入该阶段,但其活跃管线比例却明显更高,有18款药物活跃。
数据来源:药智数据、公开数据整理
百花齐放
SLE疗法迭代迅速
现阶段SLE治疗的追求并非一味要求“根治”,通常情况下寻求更合理、更低副作用的方式实现症状的“完全缓解”才是根本,因此SLE的在研新药主要围绕“激素替代”四个字进行。
而根据SLE在研新药的技术类型,目前大致可分为小分子口服药、单抗、双抗、CGT等几类。
数据来源:药智数据、公开数据整理
其中小分子药物在研管线占比最高,单抗药物发展最快,双抗与CGT疗法期望值最高,而三者也分别代表SLE治疗的过去、现在与未来。
过去:小分子口服药
小分子新药的最大优势在于其可口服、合成工艺简单、生物利用度高,因此SLE新药研发的早期大多都是围绕小分子药物进行。
发展至今,小分子SLE在研新药的构成已相当复杂,既有新一代糖皮质激素“甲泼尼龙琥珀酸钠 ”已处于注册申请阶段,也有氘可来昔替尼 、乌帕替尼、奥布替尼等小分子靶向药进入临床阶段,而总体来看,由于小分子新药选择性作用于细胞表面分子或细胞内信号转导途径中的某些信号分子来缓解病情的方式成为现实,SLE小分子在研新药也逐渐迈入了“靶向治疗”时代。
目前,SLE的靶向治疗可主要分为激酶抑制剂、钙调磷酸酶抑制剂、促细胞凋亡的抑制剂与其他四大类,其中以JAK抑制剂与BTK抑制剂为主的激酶抑制剂,在所有在研管线中占比最大。
数据来源:药智数据、公开数据整理
JAK抑制剂:为细胞内非受体型蛋白酪氨酸激酶,包括JAK1、JAK2、JAK3、TYK2四个成员,其抑制剂能够抑制Janus 激酶信号传导及转录激活蛋白(JAK-STAT)通路,影响细胞因子的信号转导,从而减少异常免疫反应,其在SLE治疗中时有新进展。研究较靠前的管线有BMS的氘可来昔替尼、艾伯维的乌帕替尼、礼来的巴瑞替尼;国内企业相关管线则有石药欧意的SYHX-1901、爱科诺生物的AC-201、恒瑞医药的硫酸艾玛昔替尼等。
BTK抑制剂:为B细胞受体信号通路中的关键激酶,生理情况下,BTK参与调节B细胞成熟、增殖、分化、凋亡、细胞迁移等过程。其抑制剂能够减少B细胞及固有免疫细胞的活化,进而调控异常免疫反应。该抑制剂也是SLE在研小分子靶向药物中的主要方向,目前研究靠前主要有诺诚健华的奥布替尼、基因泰克的非卢替尼、默克的依伏替尼、BMS的伯来布替尼。
现在:单抗类药物
在越来越多SLE发病机制的关键通路被揭示的过程中,除了小分子靶向制剂发展迅速外,靶向单抗也是目前最受期待的创新疗法之一。
其中B、T淋巴细胞受体与细胞因子信号通路在SLE抗体药物研发应用中最频繁,是目前靶向治疗研究的最主要方向,目前针对这些关键细胞/通路开发的单抗类药物约30余种,比如熟知的贝利尤单抗、泰它西普、阿尼鲁单抗等,另外还有多种靶向单抗已取得突破性成果。
表1. 部分活跃SLE单抗靶向疗法及分类(点击查看大图)
数据来源:药智数据、公开数据整理
从数据上来看,生物制剂在SLE 治疗中的地位正在逐渐提高,其作用直接、副作用更小等特点也导致靶向抗体药物等生物制剂在系统性红斑狼疮治疗中发挥着越来越重要的作用。细节上,在靶向单抗领域,靶向B淋巴细胞方向的在研药物数量明显高于企业几者,这主要源于B细胞更直接参与SLE系统性器官损伤的自身免疫致病过程,是引起自身抗体产生和免疫复合物在脏器沉积、激活补体导致器官功能进行性损伤的重要原因。
不过,现阶段的靶向抗体药物并非已经足够完美,其在有效性方面仍存在不少问题,多数只能短时间内缓解患者的症状,因此SLE靶向单抗药物的主要研究方向仍是寻找疗效更显著、更持久的方法。此外,由于SLE存在多种细胞/通路异常且交互关联,未来多靶点联合治疗、治疗性抗体的细胞内递送突破或可为领域带来新的机遇。
未来:双抗、CGT
现如今,双抗在自免领域的研究越来越受关注,目前已取得不少前期成果,靶向B细胞和浆细胞是治疗多种自身免疫疾病的关键策略,相较上述靶向单抗药物而言,其具有更强特异性结合、更低不良反应、更低药物剂量、更性价比生产成本等多种优势。
目前来看,双抗成为继单抗药物之后,SLE患者全新的治疗选择。
据不完全统计,目前全球活跃的SLE双/三抗在研管线数量不多,也尚无任何一款治疗系统性红斑狼疮的双/三抗获批上市,目前SLE双抗疗法的最高临床阶段仅位于临床II期,预计短期内仍将处于来持续探索阶段。
表2.部分“活跃”SLE双/三特异性抗体在研管线(点击查看大图)
数据来源:药智数据、公开数据整理
其中,贝林妥欧单抗与MGD-010两款新药的临床进展最快,均处于临床II期,前者作为全球第一个批准的双特异性T细胞衔接器药物,自2014年获得FDA批准用于治疗复发/难治性急性淋巴细胞白血病以来,其临床应用逐渐辐射,应用指征也在不断前移。
同时,Cullinan的CD19xCD3双特异性抗体CLN-978用于治疗中度至重度系统性红斑狼疮(SLE)患者的适应症近日也获FDA新药临床试验(IND)申请,也是首个在自身免疫性疾病中获得FDA批准IND的CD19 T细胞衔接器(TCE)。
另外,罗氏也拥有一款CD20/CD3双抗Mosunetuzumab,同时结合T细胞上的CD3和B细胞上的CD20,诱导T细胞靶向杀伤肿瘤细胞。其已经被纳入针对红斑性狼疮的1期临床试验,以评估其在该疾病中的安全性和有效性。该试验旨在评估Mosunetuzumab在SLE患者中的递增剂量的安全性和耐受性、药代动力学和药效学,并为后续研究的剂量选择提供依据。
除了双抗创新疗法,SLE新药的另一个未来趋势来源于CAR-T疗法。
早在2017年,首个CAR-T疗法获批用于治疗血液癌症之后,其关于系统性红斑狼疮等自身免疫性疾病的适应症拓展就已开始。直到2022年《自然医学》发表了一篇关于CAR-T细胞疗法治疗系统性红斑狼疮的最新研究,再次激起了全球系统性红斑狼疮CAR-T细胞疗法的开发热潮。
之所以CAR-T疗法在SLE治疗领域拥有足够想象空间,其主要原因在于,研究过程中发现B细胞制备的CAR-T细胞,可以攻击患者的B细胞群体,以此减少B细胞对自身正常细胞的错误攻击,可使经治疗的患者B细胞水平迅速降至接近零,有害的自身抗核抗体急剧下降,这就表明靶向CD19阳性B细胞的CAR-T细胞对治疗系统性红斑狼疮是安全的,且可能具有优于目前已有疗法的疗效。
据不完全统计,全球已有超过20项关于SLE适应证的CAR-T疗法在研,目前最高适应症状态为临床II期,共有9个创新疗法,不仅有诺华、阿斯利康等全球医药巨头参与其中,更有药明巨诺、亘喜生物、明聚生物等国内创新biotech企业领衔,充分显示了全球生物医药公司对该疗法的巨大兴趣和信心。
表3.部分“活跃”SLE领域CAR-T在研管线
药品名称
类型
靶点
原研单位
适应症最高阶段
瑞卡基奥仑赛 Rapcabtagene Autoleucel
自体
CD19
诺华制药
临床Ⅱ期
Anti-CD19 CAR-T cells,高博医院
-
CD19
北京高博医院有限公司
临床Ⅱ期
瑞基奥仑赛 relmacabtagene autoleucel(中国上市)
自体
CD19
上海药明巨诺生物科技有限公司
临床Ⅱ期
IMPT-514
-
CD19;CD20
ImmPACT Bio USA Inc
临床Ⅱ期
GC-012F
自体
CD19;TNFRSF17
亘喜生物科技(上海)有限公司;阿斯利康
临床Ⅱ期
CABA-201
-
CD19
Cabaletta Bio
临床Ⅱ期
Relma-cel
-
CD19
上海明聚生物科技有限公司
临床Ⅱ期
MB-CART19.1
自体
CD19
Miltenyi Biotec GmbH
临床Ⅱ期
Descartes-08
自体
TNFRSF17
Cartesian Therapeutics
临床Ⅱ期
数据来源:药智数据、公开数据整理
而更值得注意的是,这些疗法中大多是靶向CD19阳性B细胞,少数靶向BCMA(B细胞成熟抗原)阳性的浆细胞,并且部分企业已经开始涉及同种异体通用型CAR-T,比如邦耀生物的BRL-301、瑞加美生物的RJMty19、恩瑞恺诺生物的KN-5501、Nkarta的NKX-019等。
表4.部分“活跃”SLE领域异体CAR-T在研管线
药品名称
类型
靶点
原研单位
适应症最高阶段
RJMty19
通用型
CD19
浙江瑞加美生物科技有限公司
临床Ⅰ期
BRL-301
同种异体细胞
CD19
上海邦耀生物科技有限公司
临床Ⅰ期
NKX-019
同种异体细胞
CD19
Nkarta Therapeutics Inc
临床Ⅰ期
KN-5501
同种异体细胞
CD19
南京恩瑞恺诺生物技术有限公司
临床Ⅰ期
CNTY-101
同种异体细胞
CD19
Century Therapeutics
临床Ⅰ期
数据来源:药智数据、公开数据整理
不过,对于CAR-T疗法应用于SLE治疗方面来讲,通过异体技术解决患者可及性问题仅仅只是第一步,鉴于CAR - T细胞疗法在癌症患者身上有严重的副作用,导致致命的细胞因子释放综合征和神经毒性,因此系统性红斑狼疮的CAR-T细胞疗法副作用也是各个临床试验首先要关注的问题。从目前的早期临床试验结果来看,接受CAR - T细胞输注治疗的系统性红斑狼疮患者几乎没有出现细胞因子释放综合征、神经毒性或感染等严重副作用。
但仍不可否认,CAR-T疗法极有希望成为人类攻克SLE的关键点,也将是未来系统性红斑狼疮患者的重要治疗选择之一。
结语
显而易见,在面对SLE这一巨大的蓝海市场,海内外制药企业均在加速推进。
随着近年来越来越多关键通路被揭示,理想中的“激素替代”成为现实,靶向小分子药物、单抗、双抗与CAR-T疗法的出现也为患者提供了更多治疗期望。
并且,在SLE领域新药研究这个领域,明显可以看到更多中国biotech企业参与其中,且部分创新疗法的临床研究进度明显处于地位。
小分子口服药方面,虽然其中BMS、艾伯维、礼来、默克、辉瑞等MNC企业的布局较深,但同样不乏诺诚健华、石药欧意、正大天晴、恒瑞医药等制药强企深耕其中。
在单抗领域,其主要市场格局与小分子口服药相类似,仍多以MNC为研发中坚力量,但参与其中的国产企业性质却有所不同,多以天广实生物、康诺亚生物、智翔金泰、华博生物等创新biotech企业为主。
而在双/多抗与CAR-T领域,其市场格局却大有不同,MNC企业的集中程度大幅下降,且研发进度不再处于绝对的领导地位,反而是神州细胞、亿一生物、宜明昂科、药明巨诺、亘喜生物与明聚生物等biotech企业研发占领“高地”。
而很明显,以目前国内SLE新药研发的趋势来看,小分子、单抗等领域已不再是唯一的选择,中国biotech力量反而更集中于双/多抗、CAR-T等更新颖的市场领域。
未来,中国创新企业或真能借SLE如今“八仙过海,各显神通”趋势,完成对海外药企真正意义上的“赶超”。
来源 | 博药(药智网获取授权转载)
撰稿 | 头孢
责任编辑 | 八角
声明:本文系药智网转载内容,图片、文字版权归原作者所有,转载目的在于传递更多信息,并不代表本平台观点。如涉及作品内容、版权和其它问题,请在本平台留言,我们将在第一时间删除。
商务合作 | 王存星 19922864877(同微信)
阅读原文,是受欢迎的文章哦
细胞疗法免疫疗法并购临床申请
2024-11-21
·医药笔记
2024年11月14-19日,美国风湿病学会年会(ACR)在华盛顿召开,恩瑞恺诺CD19 CAR-NK疗法治疗复发性系统性红斑狼疮的最新临床数据公开发表。20例活动性重度复发性系统性红斑缆车患者接受了3个剂量的CD19 CAR-NK治疗。第一个级联组1例,第二个剂量组1例,第三个剂量组5例,第四个级联组13例。安全性方面,只观察到2例患者出现1级CRS(短暂发热),没有其他副作用和感染事件。有效性方面,对于8例随访6个月以上的患者,4例(50%)达到DORIS缓解,6例(75%)达到LLDAS(低狼疮疾病活动度)。
11月18日,CAR-NK先驱Fate Therapeutics宣布其通用CAR-T疗法FT819治疗受理狼疮患者的临床数据,该患者没有观察到3级及以上副作用,达到DORIS缓解,脱离所有免疫抑制剂治疗,重建B细胞主要是Naive、non-class switched的B细胞。
今年7月,邦耀生物在Cell期刊上发表了其通用CD19 CAR-T疗法治疗自身免疫病的临床数据。此次恩瑞恺诺发表了CD19 CAR-NK重度狼疮患者的20例临床数据,CAR-NK先驱Fate Therapeutics也宣布通用CAR-T治疗首例狼疮患者的优异疗效数据。通用CAR-T或CAR-NK或将在自免领域率先实现临床突破,期待其早日上市。
内容来源于网络,如有侵权,请联系删除。

细胞疗法抗体药物偶联物免疫疗法
100 项与 深圳恩瑞恺诺生物技术有限公司 相关的药物交易
登录后查看更多信息
100 项与 深圳恩瑞恺诺生物技术有限公司 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2025年07月18日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
临床前
3
2
临床1期
其他
4
登录后查看更多信息
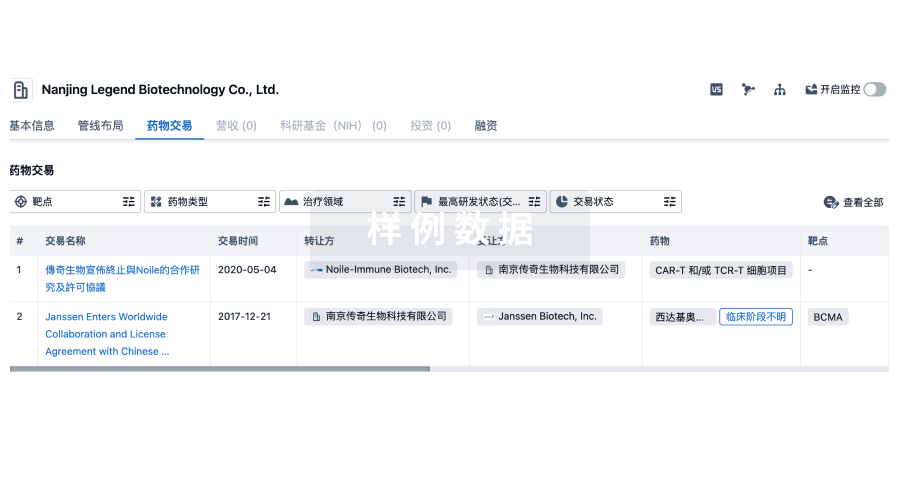
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
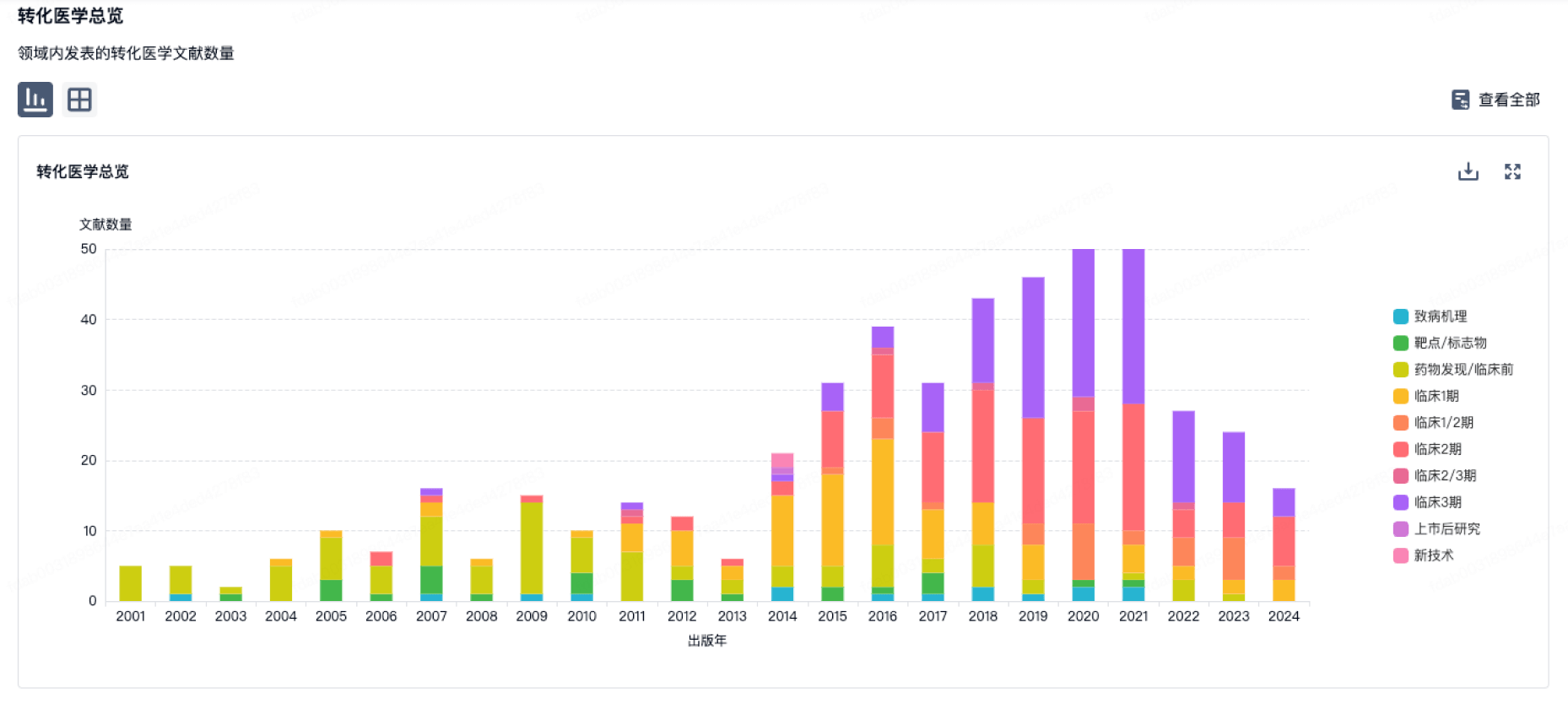
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
Eureka LS:
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用