预约演示
更新于:2025-08-29
Institute of Organic Chemistry & Biochemistry AS CR v.v.i
更新于:2025-08-29
概览
标签
感染
皮肤和肌肉骨骼疾病
免疫系统疾病
小分子化药
疾病领域得分
一眼洞穿机构专注的疾病领域

暂无数据
技术平台
公司药物应用最多的技术
暂无数据
靶点
公司最常开发的靶点
暂无数据
| 排名前五的药物类型 | 数量 |
|---|---|
| 小分子化药 | 12 |
关联
12
项与 Institute of Organic Chemistry & Biochemistry AS CR v.v.i 相关的药物作用机制 IFNβ抑制剂 [+4] |
在研适应症 |
非在研适应症- |
最高研发阶段临床1期 |
首次获批国家/地区- |
首次获批日期- |
靶点- |
作用机制- |
在研适应症 |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |

靶点 |
作用机制 IFNγ抑制剂 |
在研适应症 |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |
100 项与 Institute of Organic Chemistry & Biochemistry AS CR v.v.i 相关的临床结果
登录后查看更多信息
0 项与 Institute of Organic Chemistry & Biochemistry AS CR v.v.i 相关的专利(医药)
登录后查看更多信息
15
项与 Institute of Organic Chemistry & Biochemistry AS CR v.v.i 相关的新闻(医药)2025-08-25
撰文 | eternallj
2025年8月20日,斯坦福大学医学院Kacper B. Rogala和捷克科学院有机化学和生物化学所David M. Sabatini合作在Nature 在线发表了文章Structural basis for the dynamic regulation of mTORC1 by amino acids。该文章聚焦于营养信号调控中的核心复合物 mTORC1。作为细胞生长和代谢的关键开关,mTORC1 的活性受到氨基酸的精细调节。然而,氨基酸传感器究竟如何通过 GATOR2 将信号传递给 mTORC1,一直缺乏直接的结构证据。该研究通过冷冻电镜、突变体验证和分子模拟等手段,系统地揭示了氨基酸信号传递给 mTORC1的分子基础。
研究团队首先解决了一个长期困扰的问题:GATOR2 复合物在体外不稳定,难以进行高分辨率结构解析。为此,他们设计了一个“单链版”GATOR2(sc-GATOR2),通过将核心亚基融合固定在一起,使复合物更加稳定,并能在冷冻电镜中保持完整。功能实验表明,这一构建体仍能恢复氨基酸依赖性的 mTORC1 信号,说明它在结构和功能上均具有代表性。在这一基础上,研究人员解析了 GATOR2 与两类典型氨基酸传感器——亮氨酸传感器 Sestrin2 和精氨酸传感器 CASTOR1——结合时的结构,并获得了未结合亮氨酸的 apo-Sestrin2 构象。
冷冻电镜结果显示,Sestrin2 与 WDR24 β-propeller 区域结合,而 CASTOR1 以二聚体形式跨桥结合在 MIOS β-propeller 上,两者结合位点互不重叠,保证了不同氨基酸信号能够独立传递。为了验证这些界面是否关键,研究人员进行了共免疫沉淀实验。当 WDR24 或 MIOS 上的关键残基被突变后,Sestrin2 或 CASTOR1 与 GATOR2 的结合明显减弱,相应的 mTORC1 活性调控也随之受损。这一系列实验为结构观察提供了有力支撑。
在机制层面,亮氨酸进入 Sestrin2 结合口袋后,会引起环区和转导片段的构象重排,进而形成新的 α 螺旋,改变与 GATOR2 的相互作用界面,使其从 GATOR2 上解离。通过构建Sestrin2 点突变体,研究人员发现这些关键区域的残基一旦被改变,即便加入亮氨酸也无法打破传感器与 GATOR2 的结合。对于 CASTOR1,精氨酸结合同样会引发局部构象变化:α7 和 α3 螺旋移动,带动一个“释放环”与 MIOS 残基发生空间冲突,迫使复合物解离。相应的突变实验进一步证明了这一“释放环”的必要性。
除了静态结构和功能验证,研究人员还利用分子动力学模拟,观察了氨基酸进入传感器结合口袋的动态过程。例如在 CASTOR1 中,模拟显示精氨酸在完全结合前会先停留在 α7 螺旋附近的过渡状态,这可能正是驱动远程全构效应的关键步骤。
总体来看,研究团队提出了一个统一的模型:在缺乏氨基酸时,传感器与 GATOR2 结合,稳定并“锁住”WDR24 的 β-propeller,从而抑制 mTORC1;而在氨基酸存在时,传感器因构象重排而从 GATOR2 解离,解除对 WDR24 的束缚,使 mTORC1 得以激活。
这项工作为长期困扰研究者的氨基酸感应机制提供了清晰的结构解释,也展示了结构生物学与功能实验结合的优势。未来,如果能解析更多传感器与 GATOR2 的复合物,或者进一步绘制完整的氨基酸感应网络,我们将更深入理解 mTORC1 的营养调控逻辑。这些发现不仅具有基础科学意义,也可能为相关疾病的治疗提供新的靶点。
原文链接:https://doi-org.libproxy1.nus.edu.sg/10.1038/s41586-025-09428-7
制版人: 十一
学术合作组织
(*排名不分先后)
战略合作伙伴
(*排名不分先后)
·
转载须知
【原创文章】BioArt原创文章,欢迎个人转发分享,未经允许禁止转载,所刊登的所有作品的著作权均为BioArt所拥有。BioArt保留所有法定权利,违者必究。
BioArt
Med
Plants
人才招聘
近期直播推荐
点击主页推荐活动
关注更多最新活动!
2025-08-22
——背景——
雷帕霉素作用靶点复合物1(target of rapamycin complex1,mTORC1)是真核细胞整合代谢信号,感知营养状态,调控胞内活动和细胞生长的关键信号枢纽,其失调与多种疾病相关。之前人们已了解胞内的游离氨基酸能够通过由GATOR1、KICSTOR以及另一营养感应枢纽GATOR2组成的约2MDa超级复合物GATOR调控的Rag GTP酶激活mTORC1。GATOR2通过其WD40结构域的β-螺旋桨结构与GATOR1、亮氨酸感受器Sestrin1和Sestrin2(SESN1、SESN2),以及精氨酸感受器CASTOR1相互作用,感知细胞内氨基酸水平。然而,这些感受器如何调控GATOR2,以及它们在结合各自氨基酸配体后如何从GATOR2上解离的机制尚不清楚。近日,斯坦福大学的Kacper B. Rogala和捷克科学院有机化学和生物化学所的David M. Sabatini课题组解析了与Sestrin2或CASTOR1结合的稳定化GATOR2复合物的结构,揭示了氨基酸充足状态如何通过变构机制传递至GATOR2,并引发结构变化以实现mTORC1的激活的机制。
开启下文阅读前,请带好小纸条:
GATOR=GATOR1+KICSTOR+GATOR2
GATOR2=WDR24+WDR59+MIOS+SEC13+SEH1L
——实验结果——
1.单链GATOR2的构建和结构解析
作者在之前研究中已经解析了GATOR2复合物的游离态结构,该复合物由WDR24、WDR59、MIOS、SEC13和SEH1L五个核心亚基共同表达后组装而成。然而作者在单颗粒分析过程中发现,大多数GATOR2颗粒并不完整,至少缺少一个参与和Sestrin2和CASTOR1互作的SEH1L或SEC13亚基。为此,作者设计了GATOR2的单链变体sc-GATOR2,分别将SEH1L或SEC13以内嵌的方式插入其天然的异源二聚体伙伴WDR24、MIOS或WDR59中。由于SEH1L或SEC13的β-螺旋桨结构需要互作伙伴提供三股β-折叠片来补全,这样的构建方式保证了GATOR2复合物组装的完整性。作者还通过细胞敲入实验验证了三个单链变体能够在缺失相应基因的细胞系中恢复氨基酸依赖性的mTORC1的激活,并且与分别表达GATOR2各组分效果相当。
然后作者HEK293F细胞中瞬时表达了sc-GATOR2,解析了3.47Å分辨率的sc-GATOR2的冷冻电镜结构。通过和野生型GATOR2结构比对,作者确认了sc-GATOR2和野生型结构的一致性(RMSD为1.1Å),并通过局部分辨率比对确认了SEH1L和SEC13密度的完整性。因此作者认为sc-GATOR2能够复现野生型GATOR2的功能。
图1.单链GATOR2的构建方式和sc-GATOR2, sc-GATOR2–Sestrin2, sc-GATOR2–CASTOR1复合物结构
2.GATOR2与Sestrin2的互作模式和感知亮氨酸的机制
为了获得GATOR2–Sestrin2复合物的结构,作者选择了一个无法与亮氨酸结合但是能够持续和GATOR2结合的Sestrin2突变体E451Q与sc-GATOR2共表达。然后以3.36Å的分辨率解析了sc-GATOR2–Sestrin2复合物的结构。
图2. sc-GATOR2–Sestrin2复合物结构和亮氨酸结合机制
sc-GATOR2–Sestrin2复合物结构中,Sertrin2结合在WDR24 β螺旋桨底面与SEH1L β-螺旋桨侧面形成的表面。其中,Sestrin2带负电的αC1、αC2螺旋上的Tyr349、Glu345和Asp364和WDR24 β-螺旋桨上带正电的Arg228存在互作(图2c),Sestrin2 αC3–αC4环上的Asp406、Asp407和Gln340形成一个氢键网络,稳定两个Sestrin2环,并与WDR24的Arg46和Arg121形成分子间氢键相互作用(图2d)。Sertrin2-WDR24-SEH1L三者的互作界面上广泛分布的疏水残基也为结合提供了支持,其中包括WDR24的Met273和Val274,Sestrin2的Leu191、Ile195、Leu351、Ile352、Leu355、Leu472和Leu479,以及SEH1L的Val110(图2e)。
接着,作者通过对WDR24缺失细胞回补WDR24突变体并通过免疫共沉淀实验证实了上述这些残基在GATOR2–Sestrin2互作中的重要性,尤其是WDR24的Arg46、Arg121和Arg228和Sestrin2的Asp406与Asp407对结合最为重要。基于这些结果,作者认为GATOR2–Sestrin2相互作用主要由WDR24上的碱性表面与Sestrin2上的酸性表面之间的静电相互作用驱动。
考虑到WDR24 β-螺旋桨是mTORC1激活所必需的,作者还尝试鉴定了WDR24激活mTORC1和结合Sestrin2时的结合模式差异,发现有些WDR24和Sestrin2结合的关键残基如Arg121的突变也会影响对mTORC1的激活,但有些如Arg46,Arg228则不影响。由此作者认为,GATOR2用部分重叠的界面既与Sestrin2结合,又参与mTORC1激活;而Sestrin是通过直接阻碍GATOR2与mTORC1的结合来发挥作用。
通过结合于sc-GATOR2的apo Sestrin2结构和结合亮氨酸的Sestrin2结构进行比较,作者发现,在Sestrin2与GATOR2结合时,Sestrin2的αC2–αC3环采取一种完全伸展的构象(图2j),而在亮氨酸结合时,该区域则折叠成覆盖结合口袋入口的紧凑的双链β-折叠“盖子”。而亮氨酸结合态Sestrin2中典型的αL1–αL2螺旋–环–螺旋结构在作者解析的结构中可能因柔性未被解析,所以作者认为亮氨酸结合与αC2–αC3环的收缩和连接区的重组相关。
图S2-1. 亮氨酸结合引起的结构改变
由于亮氨酸结合口袋与GATOR2结合界面位于Sestrin2的不同表面,亮氨酸诱导形成的αL1、αL2以及其连接区并不会直接与GATOR2发生冲突,因此作者认为存在亮氨酸驱动的变构重排来改变Sestrin2–GATOR2界面。在GATOR2结合状态下,Sestrin2的Asp406通过盐桥与WDR24的Arg46相互作用。同时,Asp406还与Sestrin2的Arg338侧链相互作用,使其朝向Sestrin2。而亮氨酸结合后,αL2中的Phe287会与Arg404冲突,使Arg404侧链朝向Arg338移动,而Arg338随后进一步靠近WDR24的Arg46,两者之间产生排斥作用。作者推测,这一变构级联反应通过Arg338与Arg46的排斥作用破坏了GATOR2–Sestrin2相互作用,如图S2-2。
图S2-2. 亮氨酸结合引起的解离机制
3. GATOR2与CASTOR1的互作模式和感知精氨酸的机制
作者采用类似制备Sestrin2复合物的方法,制备了sc-GATOR2–CASTOR1复合物,并解析了分辨率为3.40 Å的sc-GATOR2–CASTOR1 复合物的结构。
图3. sc-GATOR2–CASTOR1复合物结构和精氨酸结合机制
sc-GATOR2–CASTOR1复合物结构显示,CASTOR1以二聚体形式结合在GATOR2的支架区域,每个CASTOR1单体与一个MIOS β-螺旋桨的底面结合(图3a,b)。作者提到尽管也观察到CASTOR1结合在GATOR2另一侧的数据,但他们认为这是过量游离CASTOR1引起的非特异结合。
MIOS β-螺旋桨的三条连接环构成了CASTOR1的结合位点(图3 c,d)。其中,β3.2–β3.3环将其Arg137侧链伸入CASTOR1 ACT2–ACT4界面形成的缝隙,并与CASTOR1的Asp121形成盐桥,这是GATOR2–CASTOR1结合的关键相互作用(图3c)。两侧的MIOS环进一步通过氢键网络(Arg114、His136、Asp139、Asn184)维持其伸展构象。β2.4–β3.1环则通过Lys111、His112、Ala113的主链和侧链与CASTOR1的Ser258和Tyr118形成氢键。
然后,作者通过在MIOS 缺失的细胞系中进行回补实验,验证了MIOS Arg137对GATOR2–CASTOR1结合的重要性。Arg突变体MIOS的过表达不能引起如野生型一样对mTORC1的抑制作用,证明侧链精氨酸在介导分子互作中的重要性。
CASTOR1 的精氨酸结合口袋与GATOR2结合表面位于不同区域,因此作者推测精氨酸通过变构效应调节CASTOR1–GATOR2结合。精氨酸结合首先使口袋盖(β14–α7 环)压缩(图3h),进而导致α3螺旋发生约60°的弯折,α3螺旋弯折进一步带动β6–α3 环,使CASTOR1 Ala92与MIOS Arg137发生直接碰撞,从而破坏 CASTOR1–GATOR2 相互作用(图 3i)。
——总结——
GATOR2复合物是mTORC1信号通路中的关键营养感知枢纽,但氨基酸如何通过其对应感受器影响GATOR2的机制一直不清楚。本文作者很有创意地设计了sc-GATOR2的融合表达方式,成功解析了 GATOR2 与两类营养调控因子(Sestrin2与CASTOR1)结合时的结构,发现这两类蛋白结合在互不重叠的位点,并通过不同但部分共有的机制 抑制 GATOR2 功能。这为揭示 mTORC1 营养感知通路的调控逻辑的提供了重要线索,也为理性设计相关调控药物提供了理论基础。
参考文献:
Valenstein, M.L., Wranik, M., Lalgudi, P.V. et al. Structural basis for the dynamic regulation of mTORC1 by amino acids. Nature (2025). https://doi-org.libproxy1.nus.edu.sg/10.1038/s41586-025-09428-7
作者:郭 政
审稿:钟书辰
编辑:黄志贤
GoDesign
ID:Molecular_Design_Lab
( 扫描下方二维码可以订阅哦!)
细胞疗法
2025-07-14
·精准药物
(来源:药研视角公众号)近日,国际权威药物化学期刊《Journal of Medicinal Chemistry》正式宣布成立青年编辑委员会(Early Career Board,简称 ECB),这一举措标志着药物化学领域在人才培养和学术交流方面迈出了重要的一步。来源https://pubs-acs-org.libproxy1.nus.edu.sg/doi/10.1021/acs.jmedchem.5c01588《Journal of Medicinal Chemistry》作为药物化学领域备受推崇的学术期刊,自创刊以来,一直是研究人员发表化合物设计、优化以及药物递送和成像剂的生物评估等研究成果的重要平台。此次成立的青年编辑委员会,旨在为处于职业早期的科学家们提供一个积极参与编辑工作、为期刊发展贡献力量的机会,让他们能够在这个平台上分享自己的见解、专业知识,并携手合作,共同塑造药物化学的未来。青年编辑委员会汇聚了来自不同研究领域、不同文化背景以及不同地域的优秀职业青年科学家,充分体现了多样性对于推动创新的重要性。委员会成员们将参与到为期刊撰写内容、征集主题系列稿件、推广期刊内容以及撰写每期期刊特色栏目等工作中。这些青年编辑委员会成员们,不仅在各自的学术领域取得了显著成就,更展现了对推动药物化学发展的热忱与担当。他们的加入,将为《Journal of Medicinal Chemistry》注入新的活力,为全球药物化学研究的交流与合作搭建起更加坚实的桥梁,助力更多优秀的研究成果转化为实际的治疗进步,为人类健康事业做出更大的贡献。委员会成员:王帅 副研究员 复旦大学王帅,复旦大学化学系副研究员,国家高层次青年人才,博士生导师,2023上海市白玉兰浦江人才计划,担任中国药学会药物化学专业委员会第十届委员会秘书以及青年委员、亚洲药物化学联盟秘书(AFMC,中国)、上海市药学会第十届药物化学专业委员会委员以及中国药学会药物化学决策咨询专家团队青年研究员,受邀担任国际著名药物化学期刊《Journal of Medicinal Chemistry》青年编委联合主席,《Acta Pharmaceutica Sinica B》青年编委,《Chinese Chemical Letters》青年编委和《European Journal of Medicinal Chemistry》客座主编。研究领域主要集中广谱抗病毒创新药物研究和重大慢病创新药物研究。积极推动国内外的合作与交流,分享最新的研究成果,探讨行业发展趋势,共同推动我国药物化学领域的进步与创新。王博士于2021年加入复旦大学,担任助理研究员,并于2022年晋升为副研究员。确定了多个具有前景的临床前候选药物,其中一些正在进行进一步的开发。作为一名青年科学家的倡导者,他积极指导学生和博士后研究人员,培养创新和跨学科思维的文化。“我非常荣幸能够加入《药物化学杂志》青年编辑委员会(ECB),并抓住这个机会,推动该杂志作为全球药物发现基石的使命。凭借中国药学会药物化学专业委员会秘书以及亚洲药物化学联盟(AFMC)中国区秘书的双重身份,致力于跨越地理和机构的界限,提升《药物化学杂志》的影响力和知名度。通过ECB的全球网络,我们将加速将分子发现转化为世界各地每个群体都能负担得起的药物。”徐晓莉 教授 中国药科大学徐晓莉博士是中国药科大学的教授,她的研究专长是药物化学和创新药物发现。2014年,她在本校药物化学系获得博士学位,导师是尤启冬教授。此后,她在中国药科大学中药学系继续从事博士后研究。徐博士的研究主要致力于针对分子伴侣系统和免疫肿瘤系统的药物化学、药物设计和发现。她在同行评审的学术期刊上发表了30篇学术论文,如《Journal of Medicinal Chemistry》《Cell Chemistry Biology》《European Journal of Medicinal Chemistry》等。她还获得了一些奖项,例如中国药学会颁发的2019年药物化学青年科学家奖。“我非常荣幸能够加入《药物化学杂志》青年编辑委员会,也很高兴能够与这个充满活力的新兴科学家团队和编辑委员会合作,推动该领域的创新。凭借我在基于结构的药物设计方面的专业知识,我希望能够为该领域带来新的视角,突出最前沿的趋势,并促进全球药物化学界的对话。”Nitin A. Patil 尼廷·帕蒂尔博士是北卡罗来纳州RTI国际药物发现中心的首席研究员,他领导的研究专注于开发针对多重耐药感染的下一代疗法。他在推动BRII-693(QPX9003)的发展中发挥了关键作用,这是一种目前正处于I期临床试验的新颖多粘菌素类抗生素。帕蒂尔博士的工作体现了他对转化科学的坚定承诺,他的研究处于合成化学、分子药理学和计算机辅助设计的交叉领域。帕蒂尔博士在澳大利亚墨尔本大学获得化学博士学位,此前他在印度NMIMS大学获得了药物化学硕士学位,在印度浦那大学获得了药学学士学位。他的学术和职业生涯跨越了印度、澳大利亚和美国的学术界、研究机构和制药行业。帕蒂尔博士致力于指导博士生和博士后研究员,营造一个协作和创新的研究环境。他还在药物化学、微生物学和抗微生物药理学领域教学。他通过一系列有影响力的出版物、众多国内外荣誉以及在编辑和同行评审活动中的积极领导,展现了他对药物化学界的持续承诺。除了实验室工作,帕蒂尔博士还积极参与全球科学界活动。他是美国化学学会(ACS)和澳大利亚皇家化学学会(RACI)等专业组织的成员。他目前担任《药物化学杂志》青年编辑委员会的联合主席,在那里他带来了深思熟虑的领导力和对科学严谨性的深刻尊重。除了为多个受人尊敬的期刊做出编辑贡献外,他还是20多个出版物的可信审稿人,以及国际资助机构的热门资助评审人和小组成员,包括JPIAMR、荷兰研究委员会和澳大利亚NHMRC。通过将学术卓越与战略领导力和全球经验相结合,帕蒂尔博士正在帮助塑造药物发现的未来。通过他的编辑服务和更广泛的科学参与,他继续倡导跨学科创新,并将分子发现转化为有意义的治疗进步。“我非常荣幸能够加入《药物化学杂志》的青年编辑委员会,并为在推动全球药物发现方面发挥领导作用的期刊做出贡献。我在三大洲的学术界和工业界都有工作经验,我致力于在不同学科和地区之间建立有意义的联系。作为联合主席,我旨在通过深思熟虑的编辑领导力和对严谨、有影响力科学的承诺来支持期刊的使命。我谦逊地拥抱这一特权,并期待与ECB的同事们紧密合作,通过合作、正直和共同的全球健康进步愿景来塑造药物化学的未来。Cyrille Kounde 西里尔·孔德在巴黎狄德罗大学获得了生物化学学士学位,在巴黎笛卡尔大学获得了药物化学硕士学位。在英国和新加坡的制药行业担任药物发现科学家数年后,他于2017年回到英国,在伦敦帝国学院埃德·泰特教授的团队中攻读博士学位,研究重点是蛋白降解靶向嵌合体(PROTACs)的条件激活和靶向递送。自2021年起,他在德国勃林格殷格翰公司工作,专注于发现治疗精神健康状况的新药。“将工业界与学术界联系起来可以加速药物发现,因为双方都有机会利用彼此的优势。我很高兴加入青年编辑委员会(ECB),并希望能够通过带来工业界的独特视角以及与ECB成员紧密合作来拓宽《药物化学杂志》所涵盖的主题范围。”Melissa Ashley Melissa Ashley目前是Deep Apple Therapeutics药物化学部门的首席科学家。2020年,她在哥伦比亚大学获得化学博士学位,导师是Tomislav Rovis教授。她的博士研究专注于开发光催化方法,用于选择性功能化伯胺衍生物。2020年11月,她加入Genentech的药物发现化学部门,在那里她跨多个治疗领域工作,主要专注于开发小分子抗癌药物。2025年4月,Melissa加入Deep Apple Therapeutics,在那里她利用计算工具和人工智能加速针对膜靶点的药物发现。“我非常荣幸能够加入《药物化学杂志》青年编辑委员会(ECB)。这个角色为支持药物发现领域的开创性科学以及在工业界和学术界推广高质量研究提供了令人兴奋的机会。我期待与科学界合作,塑造《药物化学杂志》的未来,并为其使命做出贡献。”Chiara Borsari Chiara Borsari于2017年在意大利摩德纳和雷焦艾米利亚大学获得药物化学博士学位。在攻读博士学位期间,她曾获得两项奖学金,分别在希腊雅典的希腊国家科学研究中心和美国纽约州立大学奥尔巴尼分校进行研究。2017年至2022年,她在瑞士巴塞尔大学生物医学系担任博士后研究员。2022年9月,她在米兰大学开始独立研究生涯,并于2023年10月晋升为药物化学终身助理教授。她的主要研究兴趣是开发选择性靶向激酶的新化学策略。近期研究亮点包括发现用于探究生物系统的化学探针,以及开发用于肿瘤学应用和治疗虫媒传染病的新共价抑制剂。她在肿瘤学领域的研究获得了L'Oréal-UNESCO女性科学奖、翁贝托·维罗内西基金会、吉多·贝尔卢奇基金会、SEED4IP概念验证和FIS启动基金的资助。她还是欧洲药物化学与化学生物学青年科学家网络(EFMC-YSN)的前任主席。Samuele CazzamalliSamuele Cazzamalli于2018年在瑞士苏黎世联邦理工学院获得了博士学位。他的博士研究获得了2018年苏黎世联邦理工学院年轻科学家奖,开创了非内化肿瘤靶向药物偶联物的开发,并首次比较了抗体-药物偶联物(ADCs)和小分子-药物偶联物(SMDCs),证明了SMDCs在靶向能力上的优势。完成博士学位后,Cazzamalli加入了瑞士-意大利生物技术公司Philochem,目前担任化学部门负责人,领导着一个由15名科学家组成的团队。他的工作涉及应用DNA编码化学库和尖端靶向策略,开发针对肿瘤相关抗原(如CAIX和FAP)的超高亲和力小分子配体。这些配体目前正在进行治疗和成像应用的临床试验,标志着精准肿瘤学的重大进展。他的团队还带头开发下一代SMDCs和放射性配体疗法。Samuele因其在肿瘤靶向领域的研究获得了2024年欧洲药物化学和化学生物学联合会(EFMC)颁发的青年药物化学家奖。“我非常荣幸能够被选为《药物化学杂志》青年编辑委员会的一员。这是一个令人兴奋的机会,能够与一群充满活力的青年研究人员和期刊的编辑团队一起贡献力量。凭借我在药物化学方面的经验,我希望能够突出该领域的创新研究和新兴方向,同时支持期刊服务于全球科学界的使命。”John Chu John Chu于2011年在斯克里普斯研究所获得化学博士学位,导师是M. Reza Ghadiri教授。随后,他在耶鲁大学的Alanna Schepartz教授和洛克菲勒大学的Sean F. Brady教授的指导下进行了博士后研究。2019年,他回到母校,在中国台湾大学化学系担任助理教授,开始了他的独立研究生涯。尽管天然产物启发了无数新药的设计,但Chu教授更广泛地将这些生物活性小分子视为自然的语言。他认为,研究它们的作用机制有助于科学家更好地理解生物体之间的相互作用,而化学正是这种交流的核心。Chu实验室采用了包括活性导向分离、人工智能工具、生物信息学分析、化学合成、克隆和基因操作等在内的经典和新方法。Chu实验室研究的一个共同主题是通过研究天然产物来学习和/或应用新的化学知识。最近的研究亮点包括将一种钙依赖性抗生素转化为硼依赖性抗生素,利用蛋白质结构预测来理解套索肽的生物合成,以及在非核糖体肽化学空间中识别过度采样/未充分探索的生态位。“作为《药物化学杂志》青年编辑委员会(ECB)的一员,我期待着通过汇聚来自世界各地、具有不同专业知识的科学家,来推动天然产物研究及其药物应用的发展。我坚信,多样性是科学创造力的关键要素。”Yoonji Lee Yoonji Lee是韩国首尔中央大学药学院的副教授。她的研究集中于化学信息学、结构生物学和机器学习的交叉领域,旨在开发用于药物靶标相互作用和先导化合物优化的预测模型。2013年,她在韩国首尔梨花女子大学获得分子建模和药物设计博士学位,导师是Sun Choi教授。2017年至2020年,她在位于美国德克萨斯州的UT西南医学中心生物物理系担任助理讲师,后升为讲师。2020年,她加入中央大学,现任副教授。在其职业生涯中,Lee教授一直致力于通过阐明配体-受体相互作用的计算方法来提高命中识别和先导化合物优化的效率。她还应用结构生物信息学系统地建模和注释蛋白质结构,为药物发现中至关重要的结构-功能关系提供关键见解。最近,她的工作已扩展到包括人工智能驱动的预测建模,以支持早期药物开发。“我非常荣幸能够加入《药物化学杂志》青年编辑委员会。我期待为该杂志持续的卓越表现做出贡献,并帮助提升药物化学创新方法的可见度。我特别兴奋能够跨学科分享想法,并突出能够加速药物发现实质性进展的数据驱动策略。”Pavla Perlikova Pavla Perlikova是布拉格化工大学(UCT)的助理教授。她的主要研究方向是基于天然化合物启发的骨架的药物化学。2012年,她在布拉格查尔斯大学获得博士学位,在捷克科学院有机化学和生物化学研究所(IOCB)从事研究,导师是Michal Hocek教授。她的研究课题集中在细胞毒性核苷和核苷酸。2012年至2013年,她在丹麦南丹麦大学与Jesper Wengel教授合作,作为博士后研究员,专注于修饰的寡核苷酸合成。随后,Perlikova博士回到捷克,在IOCB继续从事生物活性核苷和核苷酸的研究工作。2021年,她被任命为布拉格化工大学的助理教授和研究小组负责人。她的研究项目侧重于利用天然产物的骨架来开发新的生物活性化合物。她的主要研究焦点是针对肌动蛋白细胞骨架的化合物,这些化合物可能作为抗转移或抗迁移的药物。她最近的一项贡献是开发了结构简化的鬼笔环肽类似物,显示出抗迁移的效果。其他项目包括开发针对锥虫的抗寄生虫化合物以及针对各种疾病中的转录因子的化合物。“我很高兴能够加入《药物化学杂志》青年编辑委员会。我对于有机会塑造药物化学领域领先期刊的内容感到兴奋。我希望我能够帮助将一些迄今为止仍处于主流关注边缘的有趣研究话题带到聚光灯下。我期待与一群鼓舞人心的科学家合作,突出新兴的主题和创新的想法,这些想法可以拓宽我们领域的视野,并最终推进药物发现。”Chiara Borsari Roman C. Sarott是德国海德堡马普医学研究所的独立研究小组负责人。他的研究集中在使用化学诱导剂精确重编程疾病驱动因素,特别是利用小分子调节基因表达。Sarott博士于2021年在苏黎世联邦理工学院获得化学博士学位,期间在Erick M. Carreira教授的指导下研究大麻素受体的化学探针。随后,他在斯坦福大学Nathanael S. Gray教授的实验室担任瑞士国家科学基金会博士后研究员两年,开发了一种利用化学诱导邻近性原则重连癌基因激酶并诱导特定谱系癌细胞死亡的方法。这些成果为他共同创立的一家新的生物技术初创公司(位于美国南旧金山的Shenandoah Therapeutics)奠定了基础。“看到越来越多的小分子(有时也不那么小)正在重新定义我们应对疾病的方式,这令人兴奋不已。我非常荣幸能够与《药物化学杂志》的领导团队合作,推进该杂志在催化化学、生物学以及跨领域合作整合以改善人类生活方面发挥的关键作用。”Bryan J. Simmons Bryan J. Simmons是位于加利福尼亚州圣地亚哥的Bristol Myers Squibb的高级首席科学家,他在那里领导多个药物发现项目,涉及异双功能和分子胶降解剂。作为项目负责人,Bryan的努力促成了一个临床化合物的诞生,该化合物将在2025年底进入I期研究。在加入Celgene和BMS之前,Bryan在UCLA获得了博士学位,当时他是NSF研究生研究员,研究全合成以及镍催化和张力中间体方法。“我很高兴能成为《药物化学杂志》青年编辑委员会的一员,并希望能够突出那些可能对患者健康产生重大影响的新型治疗领域。”声明:发表/转载本文仅仅是出于传播信息的需要,并不意味着代表本公众号观点或证实其内容的真实性。据此内容作出的任何判断,后果自负。若有侵权,告知必删!长按关注本公众号 粉丝群/投稿/授权/广告等请联系公众号助手 觉得本文好看,请点这里↓
临床研究微生物疗法
100 项与 Institute of Organic Chemistry & Biochemistry AS CR v.v.i 相关的药物交易
登录后查看更多信息
100 项与 Institute of Organic Chemistry & Biochemistry AS CR v.v.i 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2025年10月11日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
临床前
12
3
其他
登录后查看更多信息
当前项目
登录后查看更多信息
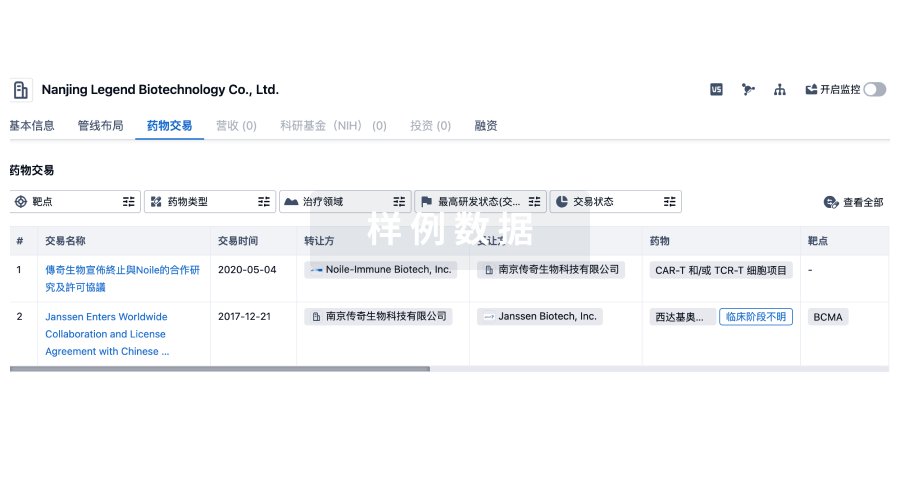
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
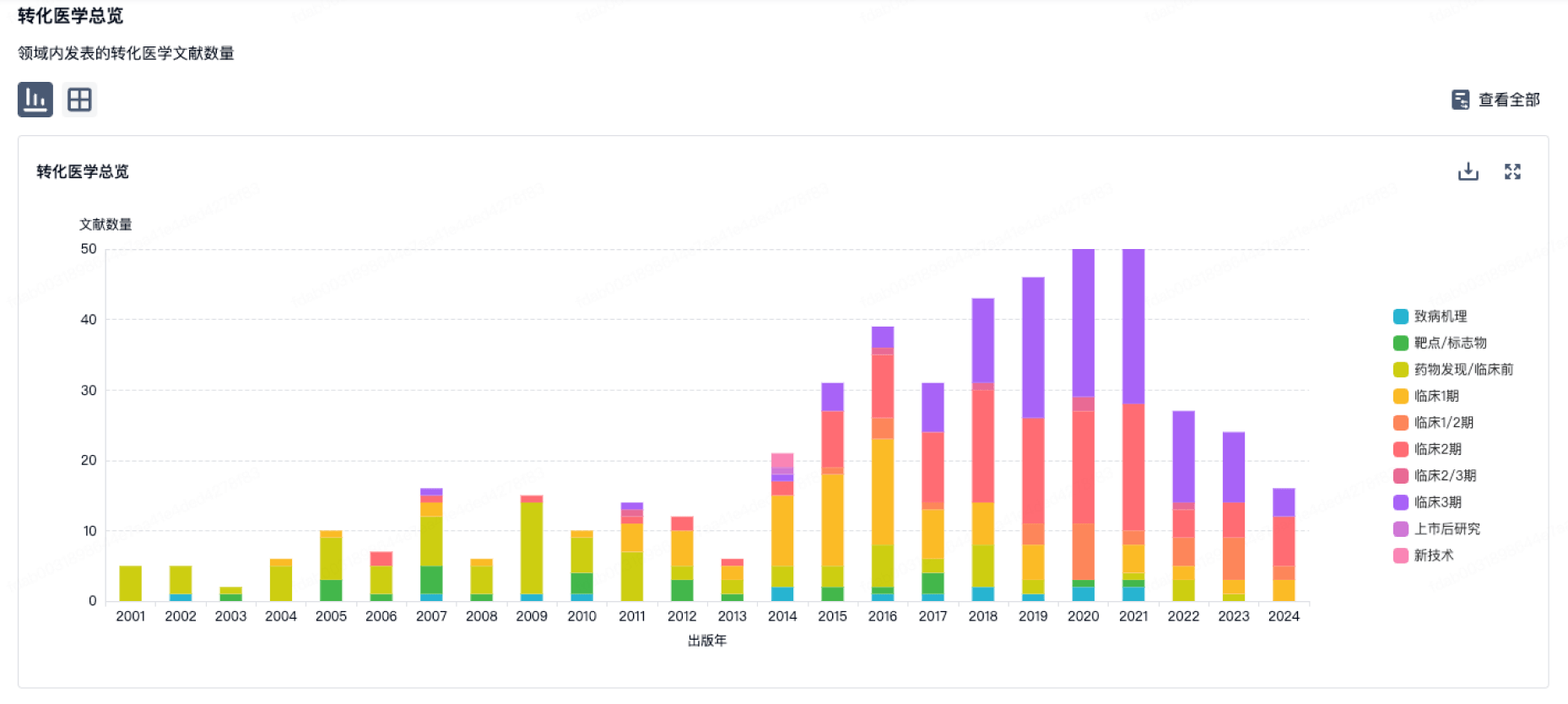
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
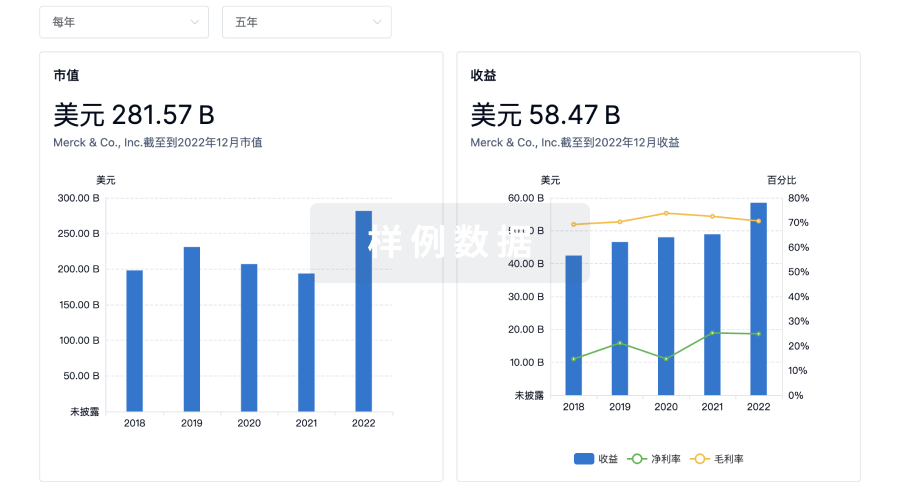
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
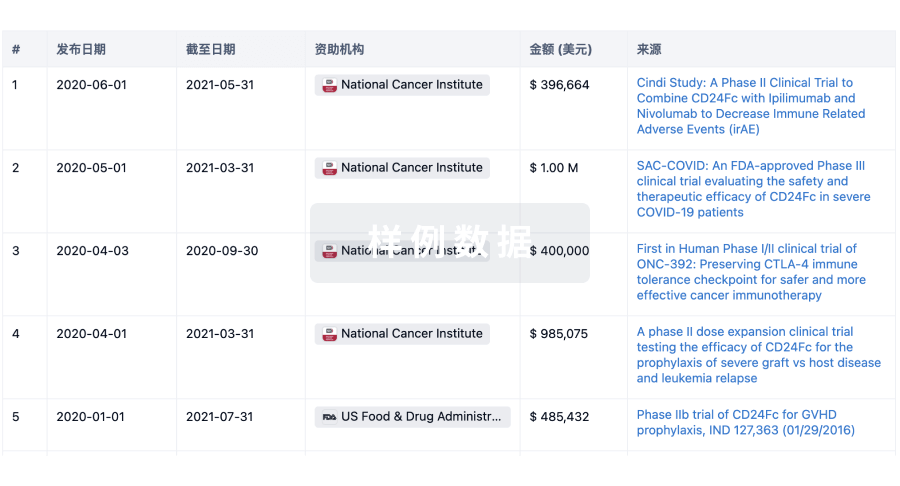
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
Eureka LS:
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用