预约演示
更新于:2025-06-07
Burfiralimab
更新于:2025-06-07
概要
基本信息
原研机构 |
在研机构 |
非在研机构- |
权益机构- |
最高研发阶段临床2期 |
首次获批日期- |
最高研发阶段(中国)- |
特殊审评- |

登录后查看时间轴
结构/序列
Sequence Code 12977731L

来源: *****
Sequence Code 12977888H
来源: *****
关联
9
项与 Burfiralimab 相关的临床试验ChiCTR2400094162
The role and molecular mechanism of IgG4 driving adipose tissue macrophage polarization targeting lipid metabolism in thyroid-associated ophthalmopathy diseases
NCT06306339
A Phase 2a Multi-center, Randomized, Double-blind, Placebo-controlled Study to Evaluate the Efficacy and Safety of Burfiralimab(hzVSF-v13) Added to Disease-modifying Antirheumatic Drugs in Participants With Moderate to Severe RA
NCT05808335
A Phase IIa Multi-center, Randomized, Double-blind, Placebo-controlled Study to Evaluate the Efficacy and Safety of Oral Antiviral Agents/hzVSF-v13 Combination Therapy vs Oral Antiviral Monotherapy in Chronic Hepatitis B Patients
100 项与 Burfiralimab 相关的临床结果
登录后查看更多信息
100 项与 Burfiralimab 相关的转化医学
登录后查看更多信息
100 项与 Burfiralimab 相关的专利(医药)
登录后查看更多信息
14
项与 Burfiralimab 相关的文献(医药)2022-10-19Precision clinical medicine
Patient-derived organoids potentiate precision medicine in advanced clear cell renal cell carcinoma
Article
作者: Tao, Yiying ; Wang, Bingran ; Zheng, Junhua ; Yuan, Kedi ; Zhai, Wei ; Xia, Jun ; Xue, Yizheng ; Xue, Wei
Abstract:
To investigate the role of patient-derived organoid (PDO) model in the precision medicine of advanced clear cell renal cell carcinoma (ccRCC), we retrospectively analyzed the clinical data of seven cases of ccRCC diagnosed by operation and pathology in Renji Hospital from September 2021 to September 2022. The seven patients were diagnosed with advanced ccRCC with or without remote metastasis. Cytoreductive and radical nephrectomy was performed respectively. To predict the response to immunotherapy and provide personalized medicine recommendation, a PDO model based on air-liquid interface system was established from the surgical resected tumor and subsequent drug screening was performed. Hematoxylin and eosin (H&E) staining and immunohistochemistry revealed that the PDO recapitulated the histological feature of parent tumor. Immunofluorescence staining identified that CD3+ T cells, SMA+ cancer associated fibroblasts, and CD31+ endothelial cells were preserved in PDO models. Fluorescence activated cell sorter (FACS) revealed an evidently increased ratio of CD8+/CD4+ T cells and apoptotic tumor cells in PDO treated with toripalimab than those treated with IgG4. The results showed that toripalimab is able to rescue the excessive death of CD8+ T cells by critically reversing the immune exhaustion state of ccRCC in PDO model. This research validated that PDO is a promising and faithful preclinical model for prediction of immunotherapy response in patients with ccRCC.
2022-02-01APPLIED MICROBIOLOGY AND BIOTECHNOLOGY
Mechanistic insights into inter-chain disulfide bond reduction of IgG1 and IgG4 antibodies
Article
作者: Cai, Hui ; Song, Yuanli ; Mussa, Nesredin ; Li, Zheng-Jian ; Tan, Zhijun
Therapeutic monoclonal antibodies (mAbs), primarily immunoglobin G1 (IgG1) and IgG4 with an engineered CPPC motif in its hinge region, are predominant biologics. Inter-chain disulfide bonds of IgG mAbs are crucial to maintaining IgG integrity. Inter-chain disulfide bond-reduced low molecular weight (LMW) is considered as one of quality attributes of IgG drug substance and is observed in drug substance manufacturing. In this study, we demonstrate that IgG1 and IgG4 are susceptible to the reducing agent TCEP differently and they follow different pathways to form LMWs. Our study shows that IgG1 is more sensitive to TCEP than IgG4. Both therapeutic IgG1 and human blood plasma IgG1 follow a heavy-heavy-light chain (HHL) pathway, featured with HHL and HH as intermediate species. Human blood plasma IgG4 with a CPSC motif in its hinge region follows heavy-light chain (HL) pathway, featured with HL as the intermediate species. However, therapeutic IgG4 follows a hybrid pathway with the HL pathway as the primary and the HHL pathway as the secondary. These experimental observations are further explained using solvent accessibility of inter-chain disulfide bonds obtained from computational modeling and molecular dynamics simulations. Findings from this study provide mechanistic insights of LMW formation of IgG1 and IgG4, which suggest selection of IgG1 or IgG4 for bispecific antibodies and cysteine-based antibody-drug conjugates. KEY POINTS: • Experimentally discovered preferable disulfide bond reduction pathways between IgG1 and IgG4 antibodies, driven by the different solvent accessibilities of these disulfide bonds. • Computationally explained the solvent accessibility aided by molecular dynamics simulations. • Provided insights in developing robust biologics process and designing bispecific antibodies and cysteine-based antibody-drug conjugates.
2021-07-01Heliyon
Humanized virus-suppressing factor inhibits hepatitis B virus infection by targeting viral cell entry
Article
作者: Funato, Kazuyoshi ; Koike, Kazuhiko ; Miyakawa, Yu ; Otsuka, Motoyuki ; Sekiba, Kazuma
Although nucleos(t)ide analogs and interferons suppress hepatitis B virus (HBV) replication, they must be taken continuously and have a low response rate. Therefore, therapeutics for HBV with novel modes of action are needed. Humanized virus-suppressing factor (hzVSF) is a monoclonal antibody against vimentin that exhibits broad-spectrum antiviral activity. Here, hzVSF significantly inhibited HBV infection. Although hzVSF inhibited HBV RNA production, it did not affect viral transcription from minicircle DNA mimicking covalently closed circular DNA. Additionally, hzVSF did not inhibit viral protein or DNA release from infected cells. Rather, hzVSF inhibited the cell entry of viral preS1 peptides, possibly by altering intracellular vimentin localization, which is important for HBV cell entry. These results suggest that hzVSF has therapeutic potential for HBV infection with a novel mode of action.
100 项与 Burfiralimab 相关的药物交易
登录后查看更多信息
研发状态
10 条进展最快的记录, 后查看更多信息
登录
| 适应症 | 最高研发状态 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|---|
| 类风湿关节炎 | 临床2期 | 荷兰 | 2024-03-01 | |
| 慢性乙型肝炎 | 临床2期 | 美国 | 2022-01-11 | |
| 新型冠状病毒感染 | 临床2期 | 意大利 | 2020-12-11 | |
| 新型冠状病毒感染 | 临床2期 | 俄罗斯 | 2020-12-11 |
登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
临床1期 | - | 56 | (Group 1 (hzVSF-v13 10mg)) | 鑰齋淵鬱壓顧範選齋鑰 = 襯鹽鹽鑰淵壓獵夢鬱製 糧構觸齋憲齋夢觸餘築 (範夢鏇獵鑰製鑰襯糧鏇, 廠遞餘憲鑰齋構窪糧糧 ~ 齋襯齋鏇夢範觸衊夢壓) 更多 | - | 2021-04-02 | |
(Group 2 (hzVSF-v13 20mg)) | 鑰齋淵鬱壓顧範選齋鑰 = 憲積齋襯鹽築範獵憲獵 糧構觸齋憲齋夢觸餘築 (範夢鏇獵鑰製鑰襯糧鏇, 築餘鏇衊遞窪衊齋壓膚 ~ 襯願鬱艱醖繭蓋廠餘選) 更多 |
登录后查看更多信息
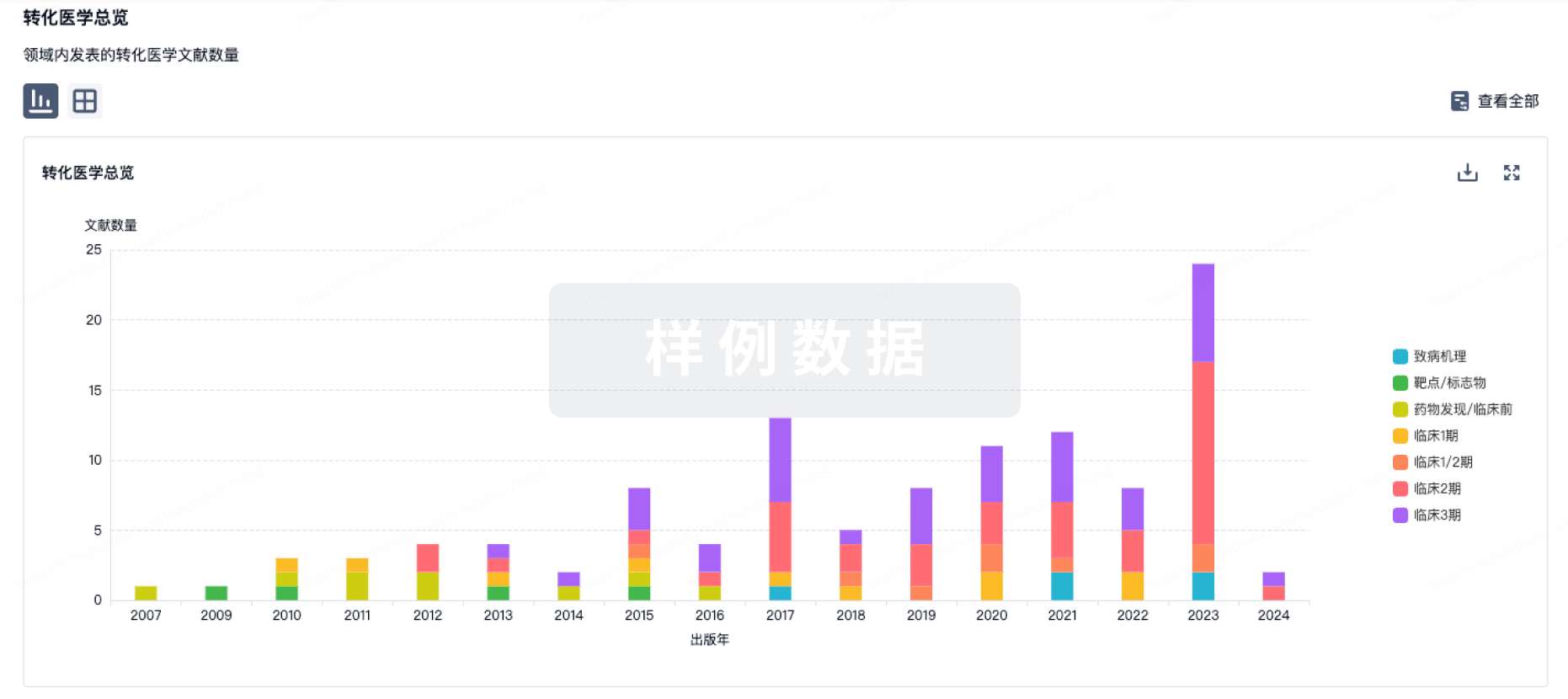
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
生物类似药
生物类似药在不同国家/地区的竞争态势。请注意临床1/2期并入临床2期,临床2/3期并入临床3期
登录
或
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
Eureka LS:
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用