更新于:2024-05-01
Sigma Systems, Inc.
更新于:2024-05-01
概览
关联
100 项与 Sigma Systems, Inc. 相关的临床结果
登录后查看更多信息
0 项与 Sigma Systems, Inc. 相关的专利(医药)
登录后查看更多信息
10
项与 Sigma Systems, Inc. 相关的新闻(医药)2024-01-20
中文摘要目的 研制以Vero细胞为生产基质的SA14-14-2乙型脑炎(乙脑)减毒活疫苗候选毒株。方法 将SA14-14-2株原始病毒在Vero细胞上传代并进行空斑克隆,对克隆株病毒进行Vero细胞培养。使用3周龄小鼠及3日龄乳鼠对脑内致病力进行测定,并进行乳鼠脑内回传、弱毒稳定性实验及免疫力实验等。结果 挑选到10个病毒克隆株,通过全面对比后,筛选到1株病毒滴度高(>6.0 lgPFU/ml)且稳定的毒株 SA14-14-2 VC6 。将该株与SA14-14-2原始株进行主要指标对比,结果表明,VC6株全基因序列与原株相比仅有1个位点发生突变(S66L),对小鼠和乳鼠的脑内致病力、弱毒稳定性、免疫原性均与原株相同。结论 筛选到的SA14-14-2 VC6株符合中国药典乙脑减毒活疫苗生产毒种的要求,各项主要指标不劣于SA14-14-2生产用毒种,是可应用Vero细胞培养生产的乙脑疫苗候选株。正文SA14-14-2乙型脑炎(乙脑)减毒活疫苗生产用细胞基质为无特定病原体(specific pathogen free,SPF)级地鼠肾细胞,SPF级地鼠群由成都生物制品研究所有限责任公司(成都生物公司)在中国首次建立(地鼠样品经英国Q-One Biotech LTD实验室全面安全检测,结果均为阴性),符合国际标准,特别是不含逆转录病毒。用地鼠肾细胞生产的SA14-14-2疫苗(SA14-4-2 PHK)于2013年通过WHO预认证,疫苗安全性已经WHO专家多次会议及大规模临床观察肯定。近20年来,随着乙脑减毒活疫苗的大量接种,乙脑发病率大幅下降,基本得到控制。尽管乙脑减毒活疫苗的安全性和保护效果已被国内外专家及WHO肯定,但个别学者认为SA14-14-2 PHK是应用动物原代细胞生产的疫苗,可能存在外源病毒污染的风险,另有报道否认WHO对应用地鼠肾细胞生产疫苗的认可,极大地影响了中国首创生产的乙脑减毒活疫苗的国际市场。另一方面,以非洲绿猴肾细胞( Vero细胞)为疫苗生产基质在生产工艺和质量控制上较地鼠肾细胞更具优势,美国已开始研制SA14-14-2株全病毒的Vero细胞减毒活疫苗。鉴于发展 SA14-14-2 Vero细胞株的重大意义,本课题组开始研发SA14-14-2株Vero细胞疫苗。本研究将SA14-14-2株病毒在Vero细胞上传代和空斑纯化,以筛选SA14-14-2适应株,并比较其与原始株SA14-14-2的致病性、病毒稳定性、免疫原性和基因特征,探讨其用作Vero细胞生产用疫苗候选株的可能性。1材料与方法1.1细胞、病毒及动物Vero细胞、BHK-21细胞(幼地鼠肾细胞)均由中国食品药品检定研究院虫媒病毒疫苗室保存;SA14-14-2减毒活疫苗,为成都生物公司的SA14-14-2 PHK株;乙脑病毒野生株P3株由中国食品药品检定研究院虫媒病毒疫苗室保存。SPF级17~19日龄(12~14 g)KM小鼠、SPF级3~5日龄KM乳鼠均由中国食品药品检定研究院实验动物中心提供,所有动物实验均由中国食品药品检定研究院实验动物福利与伦理委员会批准〔批准代码:2019(A)062〕,并按照机构指南进行。1.2主要试剂耗材DMEM(Cat.11965-092)、MEM (Cat.11965-080)培养基,胎牛血清(Cat.10091-148),0.25%胰蛋白酶(Cat.25200-072)均购自美国Gibco公司;MEM粉末培养基(Cat.M0769)、甲基纤维素粉末(Cat.M0512)购自美国Sigma公司;病毒RNA提取试剂盒(Cat.52904)购自德国QIAGEN公司;逆转录试剂盒(Cat.A5000)购自美国Promega公司。1.3病毒传代稳定性及空斑纯化将Vero细胞培养传代,待细胞长至致密单层时,以病毒感染复数为0.1的比例接种SA14-14-2株病毒,37 °C吸附1 h后,补加病毒维持液,置于37 °C细胞培养箱进行培养。待显微镜下观察到明显的细胞病变效应(cytopathic effect,CPE)时,离心收取上清病毒液。按上述操作将病毒连续传代。将Vero细胞转移至6孔细胞板进行培养,待细胞生长至致密单层时,使用病毒稀释液将病毒进行系列稀释,细胞37 °C吸附1 h后,补加1层琼脂覆盖物,置于37 °C细胞培养箱继续培养,5 d后补加第2层中性红覆盖物,48 h后挑选病毒形成的单个噬斑。将挑取的病毒克隆株在Vero细胞上进行病毒扩增传代。1.4病毒滴度测定将BHK-21细胞转移至6孔细胞板进行培养,待细胞生长至致密单层时,使用病毒稀释液将病毒进行系列稀释,细胞37 °C吸附1 h后,补加甲基纤维素溶液,置37 °C细胞培养箱继续培养,5 d后用结晶紫染色液室温染色20 min,流水冲洗、晾干后计算病毒滴度,以 lgPFU/ml表示。1.5脑内致病力及皮下致病力17~19日龄(12~14 g)的KM小鼠10只脑内注射病毒或病毒稀释液,每只0.03 ml,观察14 d,记录发病及死亡情况。17~19日龄(12~14 g)的KM小鼠10只皮下注射病毒或病毒稀释液,每只0.1 ml,同时空刺小鼠右侧脑腔破坏血脑屏障,观察14 d,记录发病及死亡情况。1.6乳鼠脑内致病力实验3~5日龄的KM乳鼠脑内注射不同稀释度病毒液,每只0.02 ml。观察小鼠14 d,记录发病及死亡情况,计算lg半数致死剂量(median lethal dose,LD50)/ml。1.7弱毒返祖实验3~5日龄的KM乳鼠10只脑内注射病毒,每只0.02 ml。每日观察小鼠,取最早发病的3只乳鼠,立即处死取鼠脑,每只乳鼠鼠脑以2 ml 病毒稀释液进行研磨,离心后收取上清液,即为鼠脑病毒研磨液。将研磨液进行系列稀释后,用12~14 g KM小鼠进行脑内致病毒力测定。1.8免疫原性实验17~19日龄(12~14 g)的KM小鼠皮下注射病毒或病毒稀释液,每只0.1 ml,共免疫 2组。免疫14 d后,对其中1组的每只小鼠进行采血,分离血清后以噬斑减少中和实验测定其中和抗体滴度;另1组小鼠以乙脑强毒株P3株进行腹腔攻毒,每只0.3 ml,同时空刺小鼠脑腔,记录14 d内发病及死亡情况。1.9噬斑减少中和实验将BHK-21细胞以2.5×105 ml -1转移至24孔细胞板培养,待细胞长至致密单层时,加入不同稀释度的免疫血清(见1.8)与1 000 PFU/ml P3强毒株的等量混合液,置于37 °C中和90 min;中和完成后,取血清病毒混合物,37 °C 吸附于细胞1 h后,补加甲基纤维素溶液,置于 37 °C细胞培养箱继续培养,5 d后使用结晶紫染色液室温染色20 min,流水冲洗待晾干后进行滴度计算,结果采用双因素方差分析和 Tukey多重比较检验,分析不同组别中和抗体滴度的差异。病毒基因测定提取不同代次病毒RNA后反转录为cDNA。使用Oligo7以SA14-14-2序列(GenBank accession No. D90195)为模板,设计7对引物,进行分段逆转录PCR扩增,委托生工生物工程(上海)股份有限公司进行引物合成及扩增片段的测序。2结果2.1SA14-14-2病毒株在Vero细胞培养传代及空斑纯化将SA14-14-2株在Vero细胞上连续传3代,从第1代开始病毒滴度已较高,为6.7 IgPFU/ml,表明Vero细胞对SA14-14-2株很敏感,继续传2代,滴度没有进一步升高,第2、3代滴度分别为7.1和6.5 lgPFU/ml。以第3代病毒收获液进行空斑纯化克隆,得到10株克隆株,分别标记为SA14-14-2 VC1 — SA14-14-2 VC10 。2.1.1 各克隆株在Vero细胞上传不同代次的病毒滴度 10株克隆株分别在Vero细胞上连续传5代,进行滴度测定,结果见表1 。各克隆株病毒在Vero细胞上传代的过程中滴度有一定波动,但是均不低于5.0 lgPFU/ml,传至第5代滴度仍维持在6.0 lgPFU/ml以上。2.1.2 各Vero细胞克隆株对3周龄小鼠的脑内和皮下致病力 将10株病毒在Vero细胞上连续传5代,测定各代病毒的小鼠脑内致病力和皮下致病力。结果显示,各代病毒对小鼠皮下感染均不致病,而脑内感染后除VC3和VC9各有4个和3个代次的小鼠死亡外,仅VC1、VC4、VC7、VC8和VC10克隆株有1个代次的小鼠死亡。其中,有3个克隆株病毒(VC2、VC5、VC6)传8代后,每代均对小鼠无致病性,维持稳定的弱毒特性。2.1.3 各Vero细胞克隆株对乳鼠脑内致病力 实验结果显示,各克隆株对乳鼠脑内致病力高低有明显差异,高者≥5.5 lgLD50/ml,低者≤ 3.6 lgLD50/ml,其中VC6株毒力最低(表2)。2.1.4 Vero细胞克隆株弱毒稳定性实验 病毒通过3~5日龄乳鼠脑内回传1代后,测定发病乳鼠脑内病毒致病力是否提高到乙脑强毒株水平,用于评价弱毒株毒力是否返祖。按中国药典2020年版三部(药典三部)中SA14-14-2乙脑减毒活疫苗规程的标准判定,毒力≥3.0 lgLD50/0.03 ml(即≥4.5 lgLD50/ml)为毒力返祖。将P5代1、2、6号Vero克隆株进行2次重复实验,返祖致病力结果均<4.5 IgLD50/ml,符合规程标准(表3 )。2.1.5 Vero细胞克隆株的免疫原性比较 选取 VC1、VC2、VC6株分别以不同病毒剂量单次皮下免疫小鼠,各2组,免疫14 d后,其中1组小鼠每只采血测定中和抗体,另1组以103 LD50 P3强毒株腹腔攻毒0.3 ml,14 d内观察小鼠死亡情况并计算保护率。实验结果(表4 )显示,3株病毒株均有良好的免疫原性。10 PFU免疫小鼠后血清阳转率均≥50%,中和抗体几何平均滴度在30~80之间,其中VC6株在不同剂量免疫的小鼠中均能产生略高于另2株的抗体水平;从保护水平看,各克隆株以100 PFU免疫小鼠的保护率均能达到70%及以上,其中VC6株免疫后小鼠保护率最高,获得了100%保护率。以上结果显示,SA14-14-2 VC6株可以在Vero细胞上稳定增殖,病毒滴度可达6.0 lgPFU/ml以上;脑内和皮下接种对3周龄小鼠均无致病力,对乳鼠脑内致病力最低(≤3.6 lgLD50/ml),同时乳鼠回传后毒力也最低(<1.0 lgLD50/ml)。免疫原性结果显示,SA14-14-2 VC6株产生了更强的抗体应答和保护效果,以下进一步将VC6株与成都生物公司生产的乙脑疫苗SA14-14-2 PHK株进行主要指标的实验比较。2.2SA14-14-2 VC6株与SA14-14-2 PHK株的主要特征比较2.2.1 脑内致病力 SA14-14-2 VC6株对 17~19日龄成鼠无脑内致病性,与SA14-14-2 PHK株一致,2株病毒对3~5日龄乳鼠的脑内致病力无明显差异(表5)。2.2.2 乳鼠脑内回传后毒力返祖 将2株病毒在乳鼠脑内回传后进行小鼠脑内毒力比较。实验显示,2株病毒的毒力返祖结果相同,均小于1.0 lgLD50/ml,符合药典三部规定(表6 )。2.2.3 免疫原性比较 依照上节所述方法,将 2株病毒以不同病毒剂量皮下免疫小鼠,各2 组。免疫14 d后,其中1组小鼠每只采血测定中和抗体,另1组以105 LD50 P3株强毒株腹腔攻毒,14 d内观察小鼠死亡情况并计算保护率(表7)。结果显示,2株病毒诱导抗体应答的水平基本相同,SA14-14-2 VC6株在低免疫剂量时的保护力高于SA14-14-2 PHK株,SA14-14-2 VC6株保护率较原始株高1倍。2.2.4 全基因序列的比较 对2株病毒进行全基因测序(表8)。结果显示,SA14-14-2 VC6在Vero细胞上传5代后,与SA14-14-2原始株相比,仅有C-292-T(S66L)1处位点发生突变,而药典三部所规定的8处包膜蛋白关键毒力位点均未发生改变,与SA14-14-2原始株相同。3讨论当前全球广泛应用的乙脑疫苗主要有3种类型:一种是中国生产的SA14-14-2株原代地鼠肾细胞制备的乙脑减毒活疫苗,另两种为基于 SA14-14-2株全病毒制备的灭活疫苗(IC50)和SA14-14-2乙脑/黄热病(JE/YF)嵌合体活疫苗JE-CV(IMOJEV),均使用Vero细胞生产。中国自主创新研制成功的SA14-14-2株乙脑减毒活疫苗由成都生物公司生产,生产用SA14-14-2株毒种由中国药品生物制品检定所建株保存。多种敏感动物脑内注射该病毒株不致病,其表型及基因型十分稳定,免疫原性良好,能诱发较强的体液免疫和细胞免疫应答。用地鼠肾细胞生产的SA14-14-2疫苗于 1989年开始在国内应用,2006年纳入国家免疫规划,在国内各省市全面接种,至今使用量已超过10亿剂次,预防效果十分显著,乙脑年发病率持续下降。2002年开始在韩国进行临床观察,之后在尼泊尔、印度等国大规模应用,至今已出口12个国家,接种量超5亿剂次。为开发以Vero细胞为基质的SA14-14-2株新一代乙脑减毒活疫苗,本研究将SA14-14-2 乙脑减毒株在Vero细胞上培养传代并空斑纯化筛选。从多株病毒培养滴度高且毒力弱的克隆株中进一步筛选得到1株病毒滴度高、神经毒力最低且免疫原性最强的SA14-14-2 VC6克隆株。与SA14-14-2原始株对比,全基因序列仅有S66L 1个位点发生突变,且该位点已被报道与Vero细胞适应性相关;其他各项指标均非劣于原始株,符合药典三部中乙脑减毒活疫苗的毒株要求,是可应用于Vero细胞生产的SA14-14-2候选毒株。以SA14-14-2株的前膜蛋白和膜蛋白与黄热病毒(yellow fever virus,YFV)17D非结构蛋白(nonstructural protein,NS)构建的 JEV/YFV嵌合体乙脑活病毒,在Vero细胞上培养制成减毒活疫苗,该疫苗已获批上市,目前正在西方国家和亚洲一些发达国家大量应用,并以Vero细胞优于地鼠肾细胞为由在亚洲扩大应用,逐步替代中国的地鼠肾细胞培养的 SA14-14-2活疫苗。本文研究的以Vero细胞培养的SA14-14-2 全病毒疫苗与国外的JEV/YFV嵌合体病毒疫苗不同,后者缺乏乙脑病毒的NS成分,而黄病毒属病毒的NS是诱生T细胞免疫应答和保护作用的主要蛋白。其中以NS1和NS3尤为重要。在亚洲和南美洲1项为期3年的长期观察中,使用相同技术构建的YFV/登革病毒(dengue virus,DENV)四价嵌合体登革热疫苗Dengvaxia被证明缺乏保护效果,且产生了极其明显的抗体依赖性增强作用。专家普遍认为其原因主要是YFV/DENV嵌合疫苗缺乏 DENV的NS,因而不能产生针对DENV的T细胞应答和抗NS1抗体的免疫保护作用,而YFV间T细胞交叉免疫效果又很弱。记忆T细胞在YFV疫苗中对保持疫苗的免疫持久性特别重要,YFV/DENV嵌合疫苗免疫原性不全面、不持久,导致若干年后产生抗体依赖性增强效应,而难以扩大应用。近期本实验室以小鼠为模型比较了中国的SA14-14-2全病毒减毒活疫苗和国外的 YFV/JEV(JE-CV)嵌合疫苗的体液和细胞免疫应答,结果显示,对于体液免疫应答,SA14-14-2疫苗各不同剂量免疫小鼠的中和抗体滴度均略高于JE-CV组免疫小鼠;对于保护效果,在低免疫剂量组中SA14-14-2疫苗明显优于JE-CV组,小鼠保护率达到80%(JE-CV 组20%);对于细胞免疫应答,SA14-14-2疫苗免疫小鼠可针对JEV-NS3多肽产生特异性 T细胞免疫应答,而JE-CV不能,前者针对JEV 细胞裂解物总抗原和全病毒抗原产生的T细胞免疫水平也强于JE-CV。结果证实,YFV全病毒能产生针对本病毒的T细胞免疫,而以异源YFV为骨架构建的嵌合体疫苗不能产生针对疫苗病毒的细胞免疫,且保护效果显著低于全病毒疫苗。尽管以Vero细胞生产的JE-CV嵌合疫苗在西方和亚洲某些发达国家广泛应用,但并不被西方国家专家普遍看好,美国NIH已着手研究 SA14-14-2株的全病毒Vero细胞疫苗,用于发展第2代SA14-14-2全病毒减毒活疫苗。总之,以Vero细胞为基质发展SA14-14-2新一代减毒活疫苗已被国际看好。本研究筛选到的SA14-14-2 VC6株在Vero细胞上培养后病毒滴度高、神经毒力低、弱毒特性稳定,并且免疫原性略高于SA14-14-2原始株,是适用于 Vero细胞生产的SA14-14-2候选毒株,在全球具有很大的应用前景。WHO十分重视以Vero细胞为疫苗生产基质制备乙脑疫苗,目前除上述JE-CV嵌合体活疫苗,用Vero细胞生产的IC50灭活疫苗已在全球广泛应用,近年来又有印度2家公司用Vero细胞生产乙脑灭活疫苗,其中1家很快得到WHO预认证。由于以Vero细胞生产的嵌合体活疫苗和灭活疫苗,对于中国以地鼠肾细胞生产的减毒活疫苗具有很大的挑战,因此研制Vero细胞 SA14-14-2疫苗十分必要和迫切。作者刘晓辉1,2 李淼1,2 刘欣玉1 徐宏山1 房恩岳1 俞永新1 李玉华11中国食品药品检定研究院虫媒病毒疫苗室,北京 102629;2长春生物制品研究所有限责任公司科研开发部,长春 130012通信作者:李玉华,Email:liyuhua@nifdc.org.cn引用本文:刘晓辉,李淼,刘欣玉,等. SA14-14-2乙型脑炎病毒Vero细胞疫苗候选株的筛选及其表型和基因型特征 [J]. 国际生物制品学杂志, 2023, 46(6): 315-322. DOI: 10.3760/cma.j.cn311962-20230306-00022识别微信二维码,添加生物制品圈小编,符合条件者即可加入生物制品微信群!请注明:姓名+研究方向!版权声明本公众号所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,不希望被转载的媒体或个人可与我们联系(cbplib@163.com),我们将立即进行删除处理。所有文章仅代表作者观点,不代表本站立场。
疫苗临床研究
2023-08-21
中文摘要目的 制备高效价兔抗新型冠状病毒血清并研究佐剂对抗体效价的影响。方法 将9只新西兰兔随机分为a、b、c组,分别皮下注射新型冠状病毒混合弗氏佐剂、氢氧化铝佐剂或单磷酰脂A佐剂进行免疫,0、28、42 d各免疫1次。每次免疫前及初次免疫后14 d于耳中央动脉采血,初次免疫后49 d经颈动脉采血,分离血清,进行中和抗体效价检测以及中和试验。结果 经过整个免疫程序后,a、b、c组均能产生高效价抗血清,且a组(6 144.00)最终产生的血清中和抗体效价高于b组(4 437.33)和c组(3 754.67);3组血清均可在中和试验中有效中和KMS2株新型冠状病毒,中和后培养细胞均未发生病变。结论 3种不同佐剂疫苗在新西兰兔体内均可产生高水平的免疫应答。正文2019年底由新型冠状病毒(severe acute respiratory syndrome coronavirus 2,SARS-CoV-2)引发的COVID-19在全世界暴发,目前疫情仍在全球传播。SARS-CoV-2属于冠状病毒科β-冠状病毒属,基因组为单股正链RNA,编码4种结构蛋白:刺突蛋白、包膜蛋白、膜糖蛋白及核衣壳蛋白[1-2]。疫苗作为一种极为有效的预防手段,是预防控制COVID-19大流行时的重要手段,而灭活疫苗是一种十分成熟的技术平台。在目前全球范围内至少有9个COVID-19全病毒灭活疫苗已进入Ⅲ期临床试验[3-9],2021年5月,由中国医学科学院医学生物学研究所生产的SARS-CoV-2灭活疫苗(Vero细胞)获批紧急使用。由于灭活疫苗在毒种选育和建库过程中可能会受外源因子的污染,因此需要对主种子批和工作种子批进行外源因子检测,在检测前需用非同源特异性血清中和病毒后再进行检测[10]。佐剂是能够增强机体对抗原免疫应答能力或调节机体产生特定类型免疫应答的物质[11]。弗氏佐剂和氢氧化铝佐剂是使用较为广泛的佐剂,新型佐剂单磷酰脂A(monophosphoryl lipid A,MPLA)是Toll样受体(Toll-like receptor,TLR)4的激动剂[12]。本研究通过在新型冠状病毒疫苗原液中添加3种不同佐剂来制备抗血清,以期找到适宜佐剂制备新型冠状病毒高效价中和血清。1材料与方法1.1疫苗原液、细胞与病毒SARS-CoV-2灭活疫苗原液、Vero细胞、指示病毒(经过标定已确定病毒滴度的SARS-CoV-2 KMS2株)均由中国医学科学院医学生物学研究所提供。1.2试剂与仪器弗氏不完全佐剂(批号:SLCB8702)和弗氏完全佐剂(批号:SLCC6223)均购自美国Sigma公司,氢氧化铝佐剂(批号:WB317588)购自美国Thermo公司,MPLA佐剂(批号:MPSU-41-03)购自美国Invitrogen公司,病毒稀释液、细胞生长液、细胞维持液及0.25%胰蛋白酶均由中国医学科学院医学生物学研究所质量检定室配制。DKB-600B水浴箱购自上海一恒科学仪器有限公司,3111 CO2培养箱购自美国Thermo公司,TS-100倒置显微镜购自日本Nikon公司。1.3实验动物无特定病原体级新西兰兔9只,雌性,体质量为2.0~2.2 kg,由昆明楚商科技有限公司提供,动物许可证号:SCXK(滇)K2018-0001。1.4免疫及抗血清制备将9只实验兔随机平均分为a、b、c组,采用皮下多点注射的方式进行免疫,均于0、28、42 d各免疫1次,0、28 d免疫剂量为300 μg/只,42 d免疫剂量为100 μg/只。a组初次免疫时将SARS-CoV-2灭活疫苗原液与等量的弗氏完全佐剂充分混匀乳化后进行免疫,后续2次加强免疫时将疫苗原液与等量的弗氏不完全佐剂充分混匀乳化后进行免疫;b、c组每次免疫时分别将等体积SARS-CoV-2灭活疫苗原液与氢氧化铝佐剂、MPLA佐剂混合均匀后进行免疫。在每次免疫前及初次免疫后14 d于耳中央动脉采血用于效价检测,初次免疫后49 d经颈动脉采血,收集血液于多个无菌小方瓶中,分离血清,进行中和抗体效价检测。1.5血清中和抗体效价检测将分离出的血清置于56 °C水浴箱中,灭活30 min。在生物安全3级实验室中用病毒稀释液将原倍血清预稀释10倍,在96孔细胞培养板第1行加入混匀的初始稀释度血清样品,每个血清样品2孔,100 μl/孔,2倍稀释至所需稀释度后吹打混匀,后吸出50 μl丢弃。在待检样品对应孔中加入50 μl浓度为100 CCID50/50 μl的病毒液,将96孔细胞培养板胶带固定后,采用样品转运箱转运至(37.0±0.5) ℃、5% CO2培养箱中和2 h。中和完成后每孔加入100 μl新鲜制备的Vero细胞悬液,混匀后将96孔细胞培养板胶带固定后采用样品转运箱转运至(37.0±0.5) ℃、5% CO2培养箱中培养7 d,于倒置显微镜下观察Vero细胞病变情况。对所用指示病毒进行回滴,通过Karber法计算指示病毒滴度,应在32~320 CCID50/50 μl范围内,将能抑制50%细胞病变的最高血清稀释度作为抗新型冠状病毒血清中和抗体效价,以稀释倍数的倒数表示。1.6中和试验将3组血清分别与KMS2株SARS-CoV-2进行中和试验,KMS2滴度均为6.50 lgCCID50/ml。将3组特异性抗血清倍比稀释,然后加入细胞培养液中进行观察是否出现细胞毒性。根据KMS2滴度和血清效价进行毒株和血清的稀释:取5 ml KMS2加入45 ml病毒稀释液中后充分混合,完成毒株10倍稀释;将特异性抗血清按照不产生细胞毒性并过量中和的原则进行稀释。将稀释后的50 ml毒株和50 ml特异性抗血清充分混合,于37 °C水浴中和2 h。中和完成后取3 ml样品加入到成长为致密单层的Vero细胞T25培养瓶中,补加9 ml细胞维持液;各组血清对照直接加入3 ml稀释后的相应组血清及9 ml细胞维持液于致密单层的Vero细胞T25培养瓶中。两者于37 ℃、5% CO2培养箱中培养14 d,第7天更换维持液1次,14 d后在倒置显微镜下观察细胞病变情况。1.7统计学分析采用SPSS 18.0统计软件对3组实验数据进行统计学分析,显著水平为ɑ=0.05,以P<0.05为差异有统计学意义。 2结果2.1血清中和抗体效价随免疫时间的变化对3组实验血清中和抗体效价检测结果如图1所示。初次免疫后,a、b、c组血清中和抗体效价在14 d内升高,14至28 d内缓慢升高,第28天a组血清中和抗体效价高于b、c组;经过第28天第2次免疫后,a组血清中和抗体效价呈缓慢升高趋势,b、c组快速升高,第42天b、c组血清中和抗体效价高于a组,经过第42天第3次免疫后,在第49天a组血清中和抗体效价升高且高于b、c组,而b、c组血清中和抗体效价有所下降。随着免疫时间的增加,a、b、c组佐剂均能刺激实验动物产生高效价抗血清。2.2血清中和抗体效价差异比较3组实验血清中和抗体效价差异比较如图2所示。初次免疫后第14天,a组血清中和抗体效价均值(832.00)高于b组(112.00)、c组(56.00)(t值:2.25、2.42,P>0.05);初次免疫后第28天,a组血清中和抗体效价(1 024.00)高于b组(448.00)(t值:2.30,P>0.05)和c组(96.00)(t值:3.70,P<0.05);初次免疫后第42天,b组(6 826.67)、c组(6 826.67)血清中和抗体效价均高于a组(1 600.00)(t值:6.29、6.29,P<0.05);初次免疫后第49天a组血清中和抗体效价(6 144.00)高于b组(4 437.33)、c组(3 754.67)(t值:1.37、1.92,P>0.05)。a、b、c组经过整个免疫程序后,均能产生高效价抗血清,且a组最终产生的血清中和抗体效价高于b、c组。2.3抗血清在中和试验中的应用效果a组血清经倍比稀释后,20倍稀释后的血清未产生细胞毒性;b组与c组血清经倍比稀释后,10倍稀释后的血清未产生细胞毒性。经过以上稀释度稀释的血清与KMS2进行中和试验,3组血清均未引起细胞病变(见表1),说明3组血清均可以在不产生细胞毒性的稀释度下完全中和KMS2。在之后的应用中,3组血清均可以用于疫苗毒种的外源因子检查。3讨论弗氏佐剂作为经典的佐剂,在实验动物制备抗血清方面应用十分广泛,但会形成难以吸收的包块,对实验动物产生较大伤害;氢氧化铝是目前中国药典中规定的唯一可用于人用疫苗的佐剂,可吸附抗原,能产生良好的免疫应答;MPLA为一种成熟的TLR4激动剂,含有MPLA的HBV和HPV疫苗已经在全球多国获得许可。本研究探讨弗氏佐剂、氢氧化铝及MLPA佐剂制备COVID-19灭活疫苗的特异性血清的效果,以期找到快速制备高效价特异性血清的合适佐剂及动物免疫程序。为了对比3种不同佐剂的效果,本实验选取SARS-CoV-2疫苗原液作为抗原,混合不同佐剂后对新西兰兔于0、28、42 d各免疫1次,并对初次免疫前及后14、28、42 d进行兔耳采血、第49天时进行颈动脉采血,收集血样后分离血清并进行抗血清效价测定。通过对抗血清效价测定结果的对比,我们可以看出,弗氏佐剂作为经典的抗血清制备佐剂,其作用明显,兔抗血清效价在初次免疫后49 d内均在上升,至49 d处死实验动物时,其抗血清效价仍然未到达顶点,这与弗氏佐剂与抗原混合乳化的过程中,含有抗原的液滴被油所包裹,形成稳定的颗粒[13-14],在注射到实验动物体内后可使抗原持续释放有关;而氢氧化铝及MPLA佐剂混合抗原进行动物初次免疫后42 d时,抗血清效价到达峰值,而后又慢慢降低,这表明使用这2个佐剂进行2次免疫后,已经可以在新西兰兔体内产生高水平的免疫应答,并可获得高效价的抗血清。本实验为了能快速、高效地制备抗血清,在最后1次免疫后7 d就进行了最后的血清分离,未考虑抗体效价在实验动物体内的长期趋势,之后的研究中可以进行长期的血清抗体效价检测,以达到更高效价获取血清抗体的目的。3种不同佐剂参与制备的抗血清,均可以在中和试验中有效中和KMS2毒株SARS-CoV-2,将在毒种检定方面起到重要作用。作者胡云光1,2 英志芳3 张玉平1,2 殷安国1,2 李亚东1,2 常亚军1,2 段男1,2 易力1,2 宋杰1,21中国医学科学院&北京协和医学院医学生物学研究所,昆明 650031;2国家药品监督管理局疫苗及生物制品质量控制与评价重点实验室,成都 611731;3中国食品药品检定研究院,国家卫生健康委员会生物技术产品检定方法及其标准化重点实验室,北京 102629胡云光和英志芳对本文有同等贡献通信作者:宋杰,Email:songjie@imbcams.com.cn引用本文:胡云光,英志芳,张玉平,等. 兔抗新型冠状病毒血清制备中佐剂对抗体效价的影响 [J]. 国际生物制品学杂志, 2023, 46(3):146-150. DOI: 10.3760/cma.j.cn311962-20220927-00063
疫苗临床3期临床结果
2023-08-04
中文摘要目的 研究绝对负压厂房对Sabin株脊髓灰质炎灭活疫苗(Sabin strain inactivated poliomyelits vaccine,sIPV)质量的影响。方法 按照批准的生产工艺在绝对负压厂房生产sIPV Ⅰ、Ⅱ和Ⅲ型单价原液各3批,配制3批sIPV三价半成品并分装成3批成品。按照企业注册标准(企标)对原液、半成品和成品进行全项检测,并将有效性指标(D抗原含量、效力试验)检测结果与相对负压厂房生产、检验合格的相对应中间产品和成品检测结果进行对比分析。对绝对负压厂房生产的Ⅰ、Ⅱ和Ⅲ型单价原液及成品进行2~8 ℃长期稳定性考察和25、37 °C加速稳定性考察,以进一步确认该厂房对sIPV质量的影响。结果 绝对负压厂房生产的单价原液中Ⅰ、Ⅱ和Ⅲ型D抗原含量分别为≥400、≥200、≥400 D抗原单位(D-antigen unit,DU)/ml,半成品中分别为48~84、54~90、82~128 DU/ml,成品中分别为24~42、27~45、41~64 DU/剂,各项检测结果均符合企标要求;供试品半成品半数有效剂量(50% effective dose,ED50)均高于参考品,且效力试验结果(供试品与参考品ED50比)均在警戒限内波动。相对负压与绝对负压厂房生产的sIPV原液、半成品、成品中Ⅰ、Ⅱ和Ⅲ型D抗原含量及半成品效力试验结果的差异无统计学意义(t值分别为-0.96、-1.69、-1.07、1.61、-0.85、0.07、0.00、-0.66、-1.04、1.93、0.83、1.31,P值均>0.05)。稳定性考察结果显示,在疫苗有效期内的各个考察时间点的所有检测结果均符合企标的要求。结论 绝对负压厂房生产的sIPV质量稳定、可控,未对疫苗质量造成影响。正文Sabin株脊髓灰质炎(脊灰)灭活疫苗(Sabin strain inactivated poliomyelitis vaccine,sIPV)由稳定的减毒脊灰疫苗株制备而成[1],是全球消灭脊灰的有效手段[2-3]。我国自主研发的sIPV于2015年在国内上市,安全性和免疫原性良好[4-8]。目前,国内sIPV生产均在相对负压厂房进行活病毒操作[9],但WHO 2015年发布的《世界卫生组织在消灭特定型别野生脊灰病毒和相继停用口服OPV后最大程度降低脊灰病毒设施相关风险的全球行动计划》(GAP Ⅲ)[10]及2019年发布的《脊髓灰质炎疫苗安全生产和质量控制指南》附件4[11],对生产脊灰病毒疫苗的设施提出了相对于环境绝对负压、安全防护和封存区的要求,以避免病毒外泄,进一步降低生物安全隐患。为确认绝对负压厂房生产的sIPV质量可控性,本研究将相对负压、绝对负压厂房生产的sIPV进行质量可比性研究[12-13],为sIPV的生产以及生物安全保障提供指导。1材料与方法1.1供试品与参考品供试品为在绝对负压厂房采用与相对负压厂房相同条件、经批准的生产工艺生产的sIPV Ⅰ型(BⅠ1—BⅠ3)、Ⅱ型(BⅡ1—BⅡ3)和Ⅲ型单价原液(BⅢ1—BⅢ3)各3批,以及采用以上单价原液生产的三价半成品(BB1—BB3)和成品(B1—B3)各3批。D抗原参考品和效力参考品均由中国医学科学院医学生物学研究所(IMBCAMS)制备,采用中国食品药品检定研究院的国家标准品进行标定。1.2可比性研究对比数据来源相对负压厂房生产、检验合格的sIPV Ⅰ型单价原液28批(PⅠ1—PⅠ28),Ⅱ型单价原液46批(PⅡ1—PⅡ46),Ⅲ型单价原液32批(PⅢ1—PⅢ32),三价半成品52批(PB1—PB52)和成品52批(P1—P52)的检测数据均由IMBCAMS提供。1.3实验动物异常毒性试验:28~35日龄无特定病原体(specific pathogen free,SPF)级ICR小鼠,18~22 g,每批产品试验使用量5只,空白对照5只;42~56日龄清洁级Hartley豚鼠,250~350 g,每批产品试验使用量2只,空白对照2只。效力试验:42~56日龄SPF级Wistar大白鼠,175~200 g,每批产品试验使用量40只,接种效力参考品40只,空白对照10只。以上实验动物均由IMBCAMS提供,实验动物生产许可证号:SCXK(滇)K2019-002。1.4主要试剂一抗、二抗、2 mol/L H2SO4、PBS和M199稀释液均由IMBCAMS制备,牛血清白蛋白(bovine serum albumin,BSA)购自北京索莱宝科技有限公司,0.05% Tween 20购自美国Sigma公司,酶标抗体购自美国Thermo Fisher公司,3,3´,5,5´-四甲基联苯胺购自美国Seracare-KPL公司,定量检测BSA ELISA试剂盒购自无锡博生医用生物技术开发有限公司,Vero细胞蛋白质残留试剂盒购自北京天坛生物制品股份有限公司,卡那霉素残留ELISA试剂盒购自北京勤邦生物技术有限公司,Hep2细胞购自美国典型培养物保藏中心。1.5方法按照企业注册标准(企标)[14]对单价原液、三价半成品和成品进行各项检定。1.5.1 D抗原含量测定 采用ELISA测定Ⅰ、Ⅱ和Ⅲ型D抗原含量。用PBS将Ⅰ、Ⅱ和Ⅲ型一抗按一定比例稀释后,加入酶标板,室温过夜。加封闭液(PBS+1% BSA)150 μl/孔,室温封闭1.5 h。用稀释液(PBS+0.5% BSA+0.05% Tween 20)将D抗原参考品及供试品稀释至适宜浓度后加入板中,100 μl/孔,阴性对照为稀释液100 μl/孔,4 °C过夜。用稀释液将Ⅰ、Ⅱ和Ⅲ型二抗稀释到适宜浓度,加入板中,100 μl/孔,室温孵育。加酶标抗体室温孵育1.5 h。加底物100 μl,室温显色10 min,加2 mol/L H2SO4终止。以450 nm为检测波长测吸收值,记录各孔吸收值,通过四参数曲线拟合法计算D抗原含量。判定标准:单价原液、半成品和成品中Ⅰ、Ⅱ、Ⅲ型D抗原含量应分别为≥400、≥200、≥400、48~84、54~90、82~128 D抗原单位(D-antigen unit,DU)/ml和24~42、27~45、41~64 DU/剂。1.5.2 鉴别试验 同D抗原含量测定,应证明含有脊灰病毒Ⅰ、Ⅱ和Ⅲ型D抗原。1.5.3 病毒灭活验证试验 采用细胞培养法进行病毒灭活验证。取含1 500剂D抗原量的样品接种到Hep2细胞,于(35.5±0.5)℃培养至21 d(原始培养),并于14 d和21 d取原始培养上清液分别盲传培养各14 d。原始培养结束后,观察细胞,并在原始细胞培养物上接种与单价病毒灭活液同型别的病毒进行攻击,攻击病毒量应为接近检测限的低剂量病毒,攻击后的细胞应出现细胞病变。原始细胞培养和2次盲传细胞培养均应无致细胞病变效应(cytopathic effect,CPE)发生,表示病毒灭活彻底,无活病毒存在。1.5.4 外观检测 肉眼观察,应为橘红色或橘黄色液体,澄清透明无异物。1.5.5 效力试验 供试品和效力参考品分别按未稀释、1:3、1:9、1:27稀释度接种Wistar大白鼠,每个稀释度接种10只,每只腿部肌内注射0.5 ml。10只Wistar大白鼠接种M199稀释液,0.5 ml/只作为阴性对照。饲养观察21 d,采血分离血清,于56 ℃灭活30 min。采用中和试验方法检测血清中的抗Ⅰ、Ⅱ和Ⅲ型脊灰病毒抗体,以1:4为阳性判定标准,结果用Reed-Muench方法计算半数有效剂量(50% effective dose,ED50),供试品的ED50应不低于效力参考品的ED50。1.5.6 装量、pH、游离甲醛、蛋白质含量、DNA残留、无菌、细菌内毒素、异常毒性、2-苯氧乙醇含量检测 按照中国药典2015年版四部通则的相关要求进行检测[15],其中,异常毒性检查采用ICR小鼠及豚鼠进行试验,2-苯氧乙醇含量采用高效液相色谱法检测。判定标准:装量应不少于0.5 ml,pH应在6.5~7.5,游离甲醛应≤25 μg/剂,蛋白质含量应≤10 μg/剂,DNA残留应≤50 pg/剂,无菌检查、异常毒性应为阴性,细菌内毒素应<10 内毒素单位/剂,2-苯氧乙醇含量应在4.0~6.0 mg/ml。1.5.7 BSA、Vero细胞蛋白质、抗生素残留检测 分别采用定量检测BSA ELISA试剂盒、Vero细胞蛋白质残留试剂盒及卡那霉素残留ELISA试剂盒进行检测,BSA残留应≤50 ng/剂,Vero细胞蛋白质残留应≤200 ng/剂,抗生素残留应≤50 ng/剂。1.6统计学分析1.6.1 趋势分析 分别计算PⅠ1—PⅠ28、PⅡ 1—PⅡ46、PⅢ1—PⅢ32、PB1—PB52、P1—P52各型别D抗原含量均值(x̅)、标准差(s)、警戒上限(x̅+2s)、警戒下限(x̅-2s)、行动上限(x̅+3s)和行动下限(x̅-3s),对BⅠ1—BⅠ3、BⅡ1—BⅡ3、B Ⅲ1—BⅢ3、BB1—BB3、B1—B3各型别 D 抗原含量进行趋势分析;分别计算PB1—PB52各型别效力试验结果的x̅、s、x̅+2s、x̅-2s、x̅+3s 和x̅-3s,对 BB1—BB3各型别效力试验结果进行趋势分析。1.6.2 独立样本t检验 使用SPSS 20.0软件分别对PⅠ1—PⅠ28和BⅠ1—BⅠ3、PⅡ1—PⅡ46和BⅡ1—BⅡ3、PⅢ1—PⅢ32和BⅢ1—BⅢ3、PB1—PB52和BB1—BB3、P1—P52 和B1—B3各型别的D抗原含量及PB1—PB52和BB1—BB3各型别效力试验结果进行独立样本t检验,P<0.05为差异有统计学意义。1.7稳定性考察对绝对负压厂房生产的sIPV Ⅰ、Ⅱ和Ⅲ型单价原液和成品各3批分别进行2~8 ℃长期稳定性考察和25、37 ℃加速稳定性考察,并绘制D抗原含量稳定性趋势图。1.7.1 单价原液 2~8 °C长期稳定性考察在0、3、6、9、12、18、24个月进行,0个月进行全项检测,3、6、9、18个月进行D抗原含量测定,12和24个月进行D抗原含量测定和无菌检查。25 ℃加速稳定性考察在0、1、2、3、4周进行,0周进行全项检测,1、2、3周进行D抗原含量测定,4周进行D抗原含量测定和无菌检查。37 ℃加速稳定性考察在0、7、14 d进行,0 d进行全项检测,7 d进行D抗原含量测定,14 d进行D抗原含量测定和无菌检查。1.7.2 成品 2~8 ℃长期稳定性考察在0、3、6、9、12、18、24个月进行,0、12和24个月进行全项检测,3、6、9、18个月进行鉴别试验及外观、蛋白质含量、2-苯氧乙醇含量、细菌内毒素、pH、D抗原含量检测。25 ℃加速稳定性考察在0、1、2、3、6个月进行,0、1、6个月进行全项检测,2、3个月进行外观、蛋白质含量、2-苯氧乙醇含量、细菌内毒素、pH、D抗原含量检测。37 ℃加速稳定性考察在0、7、14 d进行,0、14 d进行全项检测,7 d进行外观、蛋白质含量、2-苯氧乙醇含量、细菌内毒素、pH、D抗原含量检测。2结果2.1绝对负压厂房生产的sIPV检测结果2.1.1 单价原液 绝对负压厂房生产的3个型别sIPV单价原液各3批的检测结果均符合企标的要求,其中D抗原含量见表1。2.1.2 三价半成品 绝对负压厂房生产的sIPV三价半成品BB1—BB3检测结果均符合企标要求,其中D抗原含量及效力试验结果见表2。2.1.3 成品 绝对负压厂房生产的sIPV成品B1—B3检测结果均符合企标要求,其中D抗原含量见表3。2.2统计学分析2.2.1 趋势分析 相对负压厂房生产的sIPV各阶段D抗原含量及效力试验结果见表4—6。由表1一3可见,绝对负压厂房生产的sIPV Ⅰ、Ⅱ、Ⅲ型单价原液D抗原含量均在警戒限范围内波动;三价半成品Ⅰ、Ⅱ型D抗原含量在行动限范围内波动,Ⅲ型D抗原含量在警戒限范围内波动;成品Ⅰ、Ⅱ、Ⅲ型D抗原含量均在警戒限范围内波动;三价半成品Ⅰ、Ⅱ和Ⅲ型效力试验结果均在警戒限范围内波动。2.2.2 独立样本t检验 经计算,相对负压和绝对负压厂房生产的sIPV单价原液、三价半成品、成品的D抗原含量以及三价半成品Ⅰ、Ⅱ、Ⅲ型效力试验结果均服从正态分布。相对负压与绝对负压厂房生产的sIPV单价原液、三价半成品、成品中Ⅰ、Ⅱ、Ⅲ型D抗原含量差异均无统计学意义(t值分别为-0.96、-1.69、-1.07、1.61、-0.85、0.07、0.00、-0.66、-1.04,P值均>0.05);三价半成品Ⅰ、Ⅱ、Ⅲ型效力试验结果差异也均无统计学意义(t值分别为1.93、0.83、1.31,P值均>0.05)。2.3绝对负压厂房内生产的sIPV稳定性考察2.3.1 单价原液 绝对负压厂房生产的sIPV单价原液2~8 °C长期稳定性考察和25、37 °C加速稳定性考察全部时间点的所有考察项目均符合企标的要求,其中D抗原含量趋势见图1。2.3.2 成品 对绝对负压厂房生产的sIPV成品进行2~8 ℃长期稳定性考察,每一个考察时间点下的考察项目均符合企标的要求。在25 ℃、37 ℃加速稳定性考察中,除B1批Ⅰ、Ⅲ型和B3批Ⅱ型D抗原含量在25 ℃加速稳定性考察6个月的结果,以及B1、B2、B3批Ⅲ型D抗原含量在37 ℃加速稳定性考察14 d的结果低于合格下限外,其余考察项目的检测结果均符合企标的要求。sIPV成品D抗原含量考察趋势见图2。3讨论相比用野毒株生产的脊灰灭活疫苗,sIPV生产成本低,生产过程更安全,若产生泄露,所造成的生物安全风险也相对较低[10]。因此,WHO鼓励脊灰疫苗生产厂家采用Sabin株进行疫苗生产,以降低脊灰病毒传播风险,并将sIPV纳入WHO预认证高优先级别疫苗[16-18]。然而,WHO对sIPV活病毒的生产环境提出了绝对负压的要求,旨在消灭特定型别野生脊灰病毒和相继停用口服脊灰疫苗后最大程度地降低脊灰病毒设施的相关风险[10-11]。目前,全球已经进入消灭脊灰病毒的最终阶段,对sIPV需求逐年增加[19],按照WHO要求设计绝对负压厂房,生产出安全、有效的sIPV具有重大意义。本研究显示,绝对负压厂房生产的sIPV Ⅰ、Ⅱ、Ⅲ型单价原液以及三价半成品和成品各3批检测结果均合格。此外,本研究对相对负压和绝对负压厂房生产的sIPV有效性指标D抗原含量和效力试验检测结果进行了趋势分析和独立样本t检验。趋势分析结果显示,绝对负压厂房生产的sIPV各阶段D抗原含量均在行动限范围内波动,三价半成品Ⅰ、Ⅱ、Ⅲ型供试品ED50均高于参考品ED50,且两者比值均在警戒限范围内波动。经过独立样本t检验,相对负压与绝对负压厂房生产的sIPV各阶段D抗原含量、三价半成品效力试验结果差异均无统计学意义(P值均>0.05)。另外,疫苗温度标签反应速率的研究显示,2~8 ℃条件下有效期为2年的疫苗(sIPV满足该条件),可等效为25 ℃保存45 d、37 ℃保存7 d[20]。在sIPV产品有效期内的各个考察时间点的所有检测结果均符合企标的要求,绝对负压厂房生产的sIPV稳定性良好,质量可靠。绝对负压的设计可减少脊灰病毒传播,降低生物安全的风险,在今后的工作中,我们应持续关注绝对负压厂房下生产的sIPV质量,为中国及世界儿童提供安全、有效的sIPV。作者周振歆1,2 易力1,2 刘一凡1,2 杨晓蕾1,2,3 杨净思1,2,31中国医学科学院&北京协和医学院医学生物学研究所,昆明 650031;2国家药品监督管理局疫苗及生物制品质量控制与评价重点实验室,成都 611731;3病毒性传染病生物制品国家地方联合工程研究中心,昆明 650503通信作者:杨净思,Email:yjs@imbcams.com.cn引用本文:周振歆, 易力, 刘一凡, 等. 绝对负压厂房生产的Sabin株脊髓灰质炎灭活疫苗质量研究 [J]. 国际生物制品学杂志, 2023, 46(3):134-140. DOI: 10.3760/cma.j.cn311962-20221024-00072
疫苗临床结果临床3期
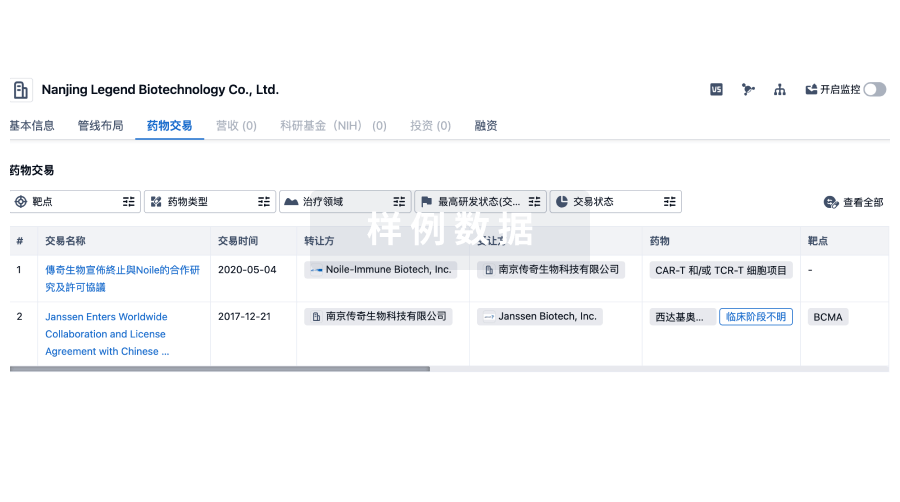
100 项与 Sigma Systems, Inc. 相关的药物交易
登录后查看更多信息
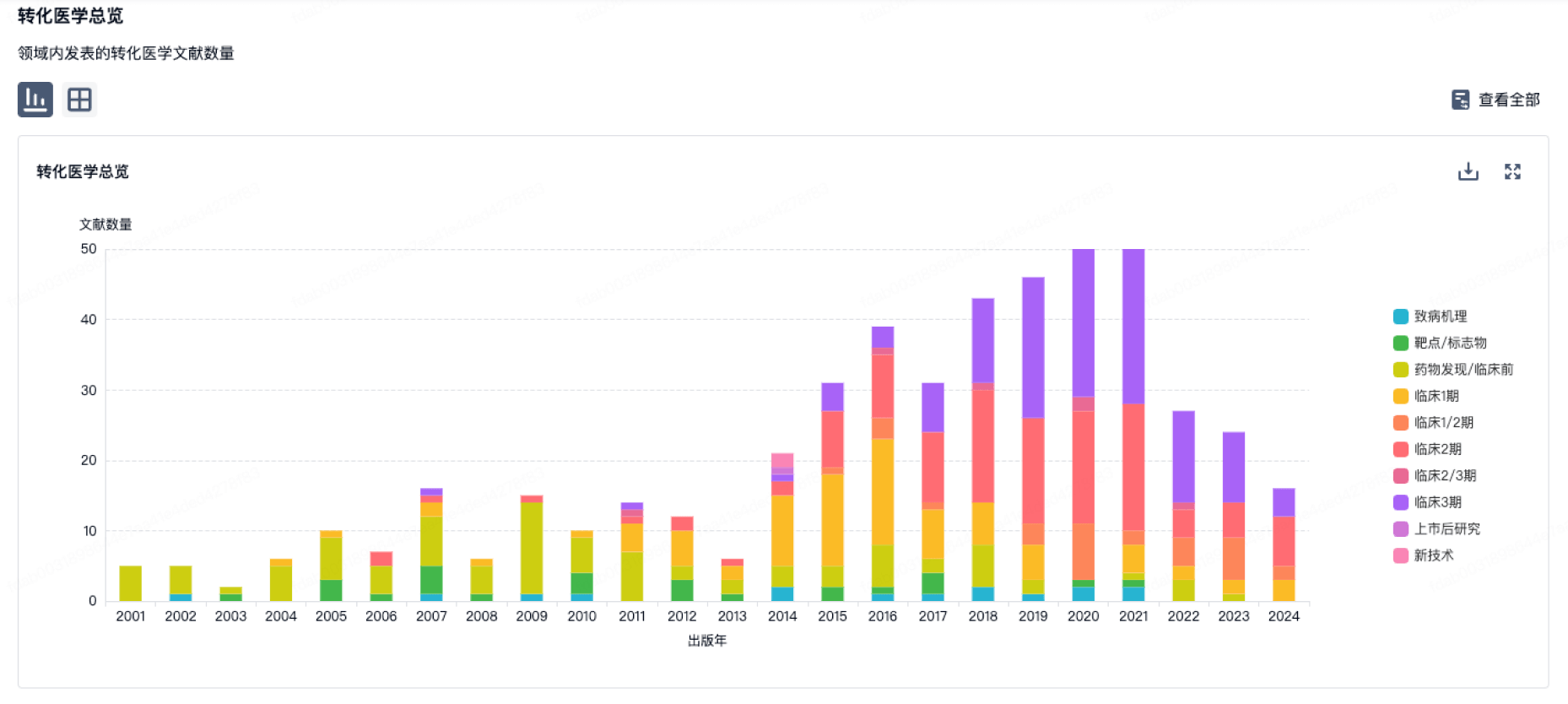
100 项与 Sigma Systems, Inc. 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2024年07月05日管线快照
无数据报导
登录后保持更新
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
标准版
¥16800
元/账号/年
新药情报库 | 省钱又好用!
立即使用
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用