更新于:2024-05-01
Milstein Dermatology LLC
更新于:2024-05-01
概览
关联
100 项与 Milstein Dermatology LLC 相关的临床结果
登录后查看更多信息
0 项与 Milstein Dermatology LLC 相关的专利(医药)
登录后查看更多信息
3
项与 Milstein Dermatology LLC 相关的新闻(医药)2024-03-26
点击上方的 行舟Drug ▲ 添加关注前言迄今为止,与传统的抗癌治疗策略相比,免疫治疗被认为是最有前景的全身性肿瘤治疗方法,其中,单克隆抗体因其特异性靶向分子的能力,已成为癌症治疗中一种关键而有效的治疗方式。然而,由于肿瘤复杂的疾病发病机制,针对单一靶点的单克隆抗体往往不足以表现出足够的治疗效果。因此,针对多个靶点的双特异性抗体(bsAbs)应运而生,它的发展改变了肿瘤免疫治疗的领域。目前,人们正在研究越来越多的针对癌症治疗的不同类型的bsAbs。这些类型包括双靶向抑制剂,其作用于不同水平的肿瘤增殖、血管形成和转移;以及双特异性抗体偶联药物、T细胞结合器、双特异性自然杀伤细胞接合器(NKCE)和先天免疫细胞接合器。其他类型还包括双特异性免疫检查点抑制剂(CPI)、共刺激bsAbs和融合蛋白、具有联合检查点抑制和共刺激特征的bsAbs、在肿瘤微环境(TME)中特异性激活的bsAb,以及将过继T细胞治疗和治疗性抗体相结合的新方法。目前,有300多项临床试验涉及200多种不同的双特异性抗体分子,其中约75%用于治疗实体瘤,25%用于治疗血液系统恶性肿瘤。有10种bsAb药物已被批准用于癌症治疗,其中9种在美国和/或欧洲获得批准,cadonilimab在中国获得批准。到目前为止,临床中开发的很大一部分bsAbs已经处于后期阶段(II期和III期)。另外,可以发现,治疗实体瘤的bsAbs主要由免疫调节剂主导,包括双CPI(约45%)和TCE(约33%),其次是靶向双途径、ICEs和ADC的bsAb。bsAb的发展史BsAbs最初的概念是由AlfredNisonof在20世纪60年代首次提出的。他将两个不同的抗原结合位点结合在一个分子中,并从抗牛γ球蛋白和抗卵清蛋白单价片段的混合物中获得了一个具有双重特异性的F(ab’)2分子。1975年,Köhler和Milstein发明了杂交瘤技术,最终解决了生产单克隆抗体的问题,开启了单克隆抗体治疗的新时代。1983年,Milstein和Cuello开创了quadroma技术,该技术基于分泌Abs混合物的两种不同杂交瘤细胞系的体细胞融合,包括具有双重特异性的鼠IgG形式的bsAb。然而,bsAb的低产率以及难以从密切相关的错配副产物中纯化所需的Ab是一个重大问题。1988年,James Huston和他的同事发明了单链抗体片段(scFv),它最大限度地减少了复性问题,例如不正确的结构域配对或聚集。1996年,Genentech的科学家们利用重组DNA技术发明了knob-into-hole技术,克服了Quadrome技术的局限性。通过在人类IgG的CH3结构域之间的界面突变选定的氨基酸,可以减少IgG重链的错配。这种技术利用更有利的蛋白质相互作用,最终可以形成高达90%的正确双特异性重链配对。随后,随着抗体工程和生物学的进展,bsAb的不同概念和结构正在不断演变。靶向双受体抑制的bsAb许多参与细胞信号传导的细胞表面蛋白,如RTK和相关受体,是基于抗体治疗的有效靶点。尽管靶向一个确定的信号受体的抗体治疗是非常有效的,但疾病相关表型通常由不止一种途径触发。这种冗余允许细胞通过使用其他补偿信号通路,从而耐受药物的生长抑制或细胞毒性诱导。同时调节不同疾病相关信号受体或通路的bsAbs可以减少或克服这一限制。Amivantamab(JNJ-61186372)靶向EGFR和肝细胞生长因子受体(MET),两种受体都能触发非小细胞肺癌(NSCLC)的增殖。因此,阻断两种受体比仅阻断一种途径能够更有效地抑制NSCLC的生长。Amivantamab已被FDA批准用于治疗携带EGFR外显子20插入突变的NSCLC亚型。一项随机III期研究的数据显示,在这部分患者中,与单独化疗相比,Amivantamab联合化疗具有显著优越性。另外一些针对肿瘤不同第二靶点的EGFR的bsAbs也正在临床开发中。例如,petosemtamab(MCLA-158)靶向EGFR和LGR5,LGR5是一种癌症干细胞相关的细胞表面受体。Petosemtamab目前正在包括晚期头颈部鳞状细胞癌在内的实体瘤患者的I/II期试验中进行评估(NCT03526835)。靶向配体受体抑制的bsAbbsAbs可以同时阻断两种不同的配体,或者是配体和受体的组合。VEGF和血管生成素2(ANG2)的双重靶向作用分别阻断VEGFR和血管生成因子1受体(TIE2)的激活,这两种途径涉及血管生成。vanucizumab,一种靶向VEGFA和ANG2的bsAb,在临床前模型中与化疗联合显示出抗肿瘤、抗血管生成和抗转移作用。BI836880是一种三特异性纳米抗体融合蛋白,由三个靶向VEGFA、ANG2和HSA的纳米抗体组成,后者可延长其血清半衰期。目前正在头颈部鳞状细胞癌、NSCLC和其他实体瘤患者的I期研究中进行评估。此外,靶向VEGF可以进一步设计成联合靶向肿瘤细胞上的PD-L1或T细胞上的PD-1,从而将抗血管生成与肿瘤免疫学方法联系起来。例如靶向PD1和VEGF的ivonescimab(AK112),其目前已在中国获批用于。其他bsAbs靶向PDL1和VEGF,包括PM8002,该药物正在NSCLC患者的II/III期研究(NCT05756972)中进行评估。靶向受体激活的bsAb许多抗肿瘤反应是通过激活细胞表面受体介导的,包括通过诱导细胞凋亡的免疫反应或细胞死亡。其中一种方法是靶向递送受体激活配体,如生长因子、细胞因子和(共)免疫刺激配体,将其融合到抗体或抗体片段中,以诱导局部或组织特异性激动活性,从而诱导细胞反应。越来越多的这种抗体-配体融合蛋白正在进入临床试验。受体激活也可以使用激动性抗体来实现。针对靶抗原和肿瘤坏死因子超家族(TNFRSF)成员的bsAbs已被证明是具有强大活性和肿瘤选择性的激动分子。RG7386(RO6874813)是一种四价bsAb,靶向癌症相关成纤维细胞上的FAPα和DR5,在具有FAPα间质表达的小鼠模型中具有效和选择性的肿瘤细胞杀伤,并已进入临床测试。此外,BI905711,一种靶向DR5和CDH17的四价双特异性IgG-scFv融合蛋白,目前处于I期临床研究中。靶向递送有效载荷的bsAb存在两种概念上不同的方法来利用bsAbs靶向递送有效载荷,如细胞毒性药物或放射性药物。第一种方法,预靶向治疗,使用bsAb的一个臂特异性靶向肿瘤细胞,另一个臂随后捕获有效载荷到肿瘤细胞。GD2 x DOTA是一种识别GD2的bsAb,并捕获放射性有效载荷177Lu-DOTA,其在TME中组装。在没有四聚化的情况下,由于半衰期,它会被迅速从外围清除。然而,到目前为止,临床试验中很少有bsAbs利用这种MoA,这可能是因为放射性预靶向涉及复杂的导流,限制了其普遍适用性。第二种方法通过递送偶联到双特异性抗体上的细胞毒性有效载荷,实现肿瘤细胞的特异性靶向。目前,bsADC已成为肿瘤药物开发的热点,已有多款bsADC进入临床开发,如ZW49、REGN5093、BL-B01D1等。靶向癌症免疫治疗的bsAb癌症免疫疗法中使用的大多数bsAbs通过T细胞驱动的天然或内源性免疫发挥作用,包括免疫检查点抑制(CPIs)效应细胞接合器(包括TCE和ICEs)以及共刺激性bsAb。双免疫检查点抑制许多抑制免疫检查点的单特异性抗体,如CTLA4或PD-1/PDL-1,已被批准用于癌症免疫疗法。将检查点抑制性抗体疗法扩展到bsAbs可以减少不期望的副作用,并增强疗效。大多数双免疫检查点抑制剂靶向PD-1/PD-L1轴和其他免疫抑制分子,如CTLA-4、TIM-3、LAG-3或TIGIT。MEDI5752(PD-1×CTLA-4)抑制PD-1+激活的T细胞中CTLA-4的功能,诱导PD-1的快速内化和降解。与PD-1和CTLA-4单抗联合治疗相比,MEDI5752的活性显著增强。针对其他检查点分子的双功能性抑制剂,包括LAG-3和TIGIT,也正在开发中。效应细胞接合器细胞接合器连接两种不同类型的细胞,主要是肿瘤细胞和T/NK细胞,以诱导肿瘤细胞溶解。细胞接合器由TAA靶向部分和效应细胞识别部分组成,并进一步分为T细胞接受器和NK细胞接受器。T细胞接合器利用TAA和TCR成分(主要是CD3),它们连接肿瘤细胞和T细胞,绕过TCR-MHC-I相互作用。因此,无论抗原特异性如何,T细胞结合器都会触发T细胞活化,产生包括穿孔素和颗粒酶在内的细胞毒性分子,以杀伤肿瘤细胞。BiTE是T细胞结合器的一种形式,通过靶向TAA和CD3的单链抗体将肿瘤细胞和T细胞结合,用于T细胞介导的肿瘤细胞杀伤。靶向CD19和CD3的blinatumomab是临床上典型的BiTE,2014年,blinatumomab成为第一个被FDA批准的BsAb,用于治疗ALL患者,在随后的几年里,blinatumomab的治疗范围进一步拓宽,2018年,FDA批准其用于首次或第二次完全缓解后MRD≥0.1%的pre-B ALL患者群体的治疗。目前,超过60例BiTE形式的双抗正在进行I/II期临床试验。NK细胞接合器将CD16A+NK细胞重定向至肿瘤细胞,诱导NK细胞活化。与BiTE相比,NK细胞接受者表现出较少的不良反应,如细胞因子释放综合征(CRS)和神经毒性。多种细胞毒性受体可激活NK细胞:CD16(FcγRIII)、天然细胞毒性受体(NCR;NKp30、NKp44、NKp46)、C型凝集素样受体NKG2D(CD314)和CD94/NKG2C。最近,抗CD30×CD16A的NK细胞接合器(AFM13)在AACR 2022的报告中展示了不俗的抗肿瘤效果。共刺激bsAbs高效、持久和局部受限的抗肿瘤免疫反应需要共刺激和共抑制信号来严格调节细胞毒性T细胞、自然杀伤细胞和巨噬细胞的激活、分化和维持。TNFRSF家族的成员(如4-1BB、CD40、OX40、GITR、CD27或CD30)以及CD28免疫球蛋白超家族(如CD28、CTLA4、PD-1、ICOS、BTLA)及其相应的配体在介导免疫调节信号中发挥着重要作用。靶向这些免疫调节途径可以增强免疫治疗。PRS-343(cinrebafusp),是一种针对HER2和4-1BB的四价bsAb,在IND研究中其耐受性良好,没有明显和相关的药物相关毒性;在接受有效剂量治疗的患者中(n=33),12%的患者达到ORR,包括1个CR,疾病控制率(DCR)为52%。bsAbs中通过4-1BB进行肿瘤选择性共刺激的其他靶点还包括EGFR、PSMA、CLDN18.2、B7H4、CEACAM5、HER2、PD-L1和FAP。此外,十多种针对PD-L1和4-1BB的不同bsAbs处于I期或II期试验中。bsAb的新概念bsAbs的领域仍在进一步探索和扩展技术和应用,可能提供目前正在开发的bsAb尚未解决的治疗概念。三特异性TCE共刺激信号可以与双特异性TCE组合以促进更加有效的T细胞活化。这样的三特异性TCE目前正在针对TAA和CD3的临床试验中,并且进一步包含抗CD28结合位点。SAR443216是一种针对HER2×CD3×CD28的三特异性抗体,能够强力激活CD4+和CD8+T细胞,诱导T细胞增殖、细胞因子和颗粒酶B的释放,以及T细胞介导的对表达HER2的肿瘤细胞的杀伤,其目前处于I期临床研究中。在类似的方法中,SAR442257是另一种CD38×CD3×CD28的三特异性抗体。前药方法一种新兴的方法是在肿瘤处选择性激活的前药,产生具有高度肿瘤特异性功能的bsAbs。前药转化为功能性bsAbs可以通过环境触发因素如肿瘤相关蛋白酶来实现。双重肿瘤靶向的另一种前药方法依赖于来自两个具有分裂CD3ε结合部分的肿瘤特异性组装。这些分子需要被设计为非活性的独立实体并在循环中,使得它们仅在肿瘤抗原结合和随后的原位组装时被激活。PROTAC bsAbs另一个新兴概念和新的MoA利用bsAbs通过靶向降解来抑制增殖相关过程。类似于双特异性或多特异性小分子蛋白水解靶向嵌合体(PROTACs),bsAbs可被设计为触发参与癌症发展或进展的表面蛋白的内化和随后的降解。这样的bsAbs同时与靶向降解的蛋白质和降解的因子结合,并触发内化,如膜上的E3泛素连接酶或转铁蛋白受体。目前,抗体-PROTAC概念仍处于早期开发阶段,迄今为止还没有进入临床研究。bsAb递送bsAb开发中另一个重要的新兴概念是bsAb的局部产生和随后的局部递送,如通过脂质纳米颗粒介导的mRNA递送的CLDN6 TCE、溶瘤病毒或包括CAR-T细胞在内的基因治疗方法。细胞因子模拟抗体最后,bsAbs也可以作为细胞因子模拟物或所谓的合成细胞因子激动剂。scFvs或基于单域抗体VHH的方法可以通过模拟IL-2或IL-15、IL-18、I型干扰素和IL-10来有效地触发细胞因子信号传导。这种基于抗体的细胞因子模拟物可能在癌症免疫疗法中具有各种应用。小结bsAbs领域最近取得了实质性进展,到2023年底,已有十几种bsAbs获得批准,许多不同的方法正在临床上进行测试。尽管如此,由于癌症的异质性和适应性,这些方法中的任何一种都不太可能成为普遍的癌症免疫疗法。相反,抗体需要针对某些应用量身定制,并将依赖于与其他方法的结合,以最大限度地提高其疗效和安全性。在不久的将来,多特异性抗体疗法的发展可能会在以下四个领域出现增长:针对具有明确依赖性的肿瘤相关RTK的bsAbs;用于以增加的选择性靶向肿瘤的bsADC;用于靶向降解适用于各种途径的细胞表面蛋白的双特异性PROTAC;以及用于癌症免疫疗法的多特异性抗体。与其他治疗癌症的药物一样,通过改进的诊断方法和新的开发模式,将bsAbs和多特异性抗体疗法应用于早期患者,对于实现治愈癌症患者的目标至关重要。参考文献:1. The present and future of bispecific antibodies for cancer therapy. Nat Rev Drug Discov.2024 Mar 6.2. Development of Bispecific Antibody for CancerImmunotherapy: Focus on T Cell Engaging Antibody. Immune Netw. 2022 Feb; 22(1): e4.文章信息源于公众号小药说药,登载该文章目的为更广泛的传递行业信息,不代表赞同其观点或对其真实性负责。文章版权归原作者及原出处所有,文章内容仅供参考。本网拥有对此声明的最终解释权,若无意侵犯版权,请联系小编删除。学如逆水行舟,不进则退;心似平原走马,易放难收。行舟Drug每日更新 欢迎订阅+医药大数据|行业动态|政策解读
免疫疗法临床2期细胞疗法临床结果引进/卖出
2022-08-04
单克隆抗体(mAb)具有高特异性并能增强免疫反应的特点,是一种潜在的病毒感染治疗剂和预防剂。而抗体工程可增强效应器功能,延长mAb半衰期。结构生物学的进步,则利于通过识别病毒蛋白中的易损区域选择和优化有效的中和mAb,并设计疫苗。COVID-19的爆发使针对严重急性呼吸综合征冠状病毒2(SARS-CoV-2)中和mAb的研发日益火热,目前已有几款mAb获得紧急使用授权(EUA)。mAb的应用不仅有益于COVID-19应对,而且还推动其他传染病的治疗和预防。在这里,我们总结了抗体发现和抗体工程的研究进展,以及抗病毒感染mAb的开发过程,这些病毒包括SARS-CoV-2、呼吸道合胞病毒(RSV)、埃博拉病毒(EBOV)、人类巨细胞病毒(HCMV)和流感病毒。自1970年代Köhler和Milstein开发能够产生单克隆抗体(mAb)的杂交瘤技术以来,mAb已成为治疗癌症和免疫疾病的关键药物,2021年第100种基于抗体的治疗药物已获得美国食品和药物管理局(FDA)批准。然而,尽管利用抗体对抗传染病的历史可以追溯到一个多世纪以前的血清应用,但只有少数mAb药物用于治疗或预防传染病。截至2022年初,已有6种针对病原体的mAb已获得FDA批准(表1),适应症包括预防呼吸道合胞病毒(RSV)感染、预防和治疗炭疽感染、预防艰难梭菌感染复发和治疗埃博拉病毒(EBOV)感染。表1:FDA批准的用于传染病适应症的mAb2020年,COVID-19的迅速传播使研究人员大力开发针对SARS-CoV-2的中和mAb。现阶段,超过20种mAb进入临床开发阶段,其中一些mAb已获得FDA和全球其他机构的EUA(表2)。尽管疫苗一直是应对COVID-19的主力军,但mAb也有一定贡献,特别是针对免疫功能低下或有轻度至中度COVID-19的患者。表2针对COVID-19上市或处于后期临床研究的mAb1抗体特性与抗体工程1抗体结构和功能抗体是病原体感染或接种疫苗后由浆细胞或受刺激的记忆B细胞产生的天然生物分子,为Y形异二聚体,由两条25kDa的轻链和两条至少50kDa的重链组成,重链和轻链之间通过多个二硫键和非共价相互作用连接(图1a)。抗体也可分为抗原结合结构域(Fab)和可结晶片段结构域(Fc)(图1b),两个Fab结合并中和病原体,通过铰链与Fc相连,Fab相对Fc具有较大的构象灵活性,能与抗原产生强烈相互作用。糖基化的Fc结构域与其他蛋白质结合,可以介导抗体依赖性细胞毒性(ADCC)、补体依赖性细胞毒性(CDC)和抗体依赖性等细胞吞噬作用(图1c)。Fc结构域还可以与新生儿Fc受体(FcRn)相互作用,影响抗体药代动力学。抗体的种类取决于alpha、mu、gamma、epsilon、delta等基因片段是否与其可变区重组。人类以下基因产生不同的抗体亚类:α基因片段产生IgA1和IgA2,γ基因片段产生IgG1、IgG2、IgG3和IgG4,mu片段产生IgM,epsilon片段产生IgE和delta片段产生IgD。除IgD外,每个亚类都能消除不同类型的病原体,IgG类主要针对血液和细胞外液中的病原体。不同抗体亚型的序列变异决定了它们对FcRn、FcγR和补体蛋白C1q的亲和力和特异性。其中IgG1启动ADCC和CDC(图1c),IgG2和IgG4是较差的CDC激活剂,而IgG3有效激活CDC,大多数临床使用或针对感染性疾病开发的治疗性mAb是IgG1亚型。图1:病毒感染期间单克隆抗体的作用机制2抗体作为抗病毒药物一个多世纪以来,单克隆或多克隆抗体已被用于传染病治疗和预防。抗体可以通过多种机制对抗病毒感染。首先,抗体可以防止包膜病毒的病毒糖蛋白或无包膜病毒的蛋白壳与靶宿主细胞结合,这些病毒蛋白在病毒生命周期中具有两个主要功能:与细胞受体结合,介导病毒和细胞膜融合(在包膜病毒的情况下)或渗透到细胞质中(在无包膜病毒的情况下)。例如,SARS-CoV-2进入宿主细胞是由病毒刺突(S)糖蛋白与宿主细胞表面的血管紧张素转换酶2(ACE2)受体之间相互作用介导的。ACE2在呼吸系统、胃肠道和内皮细胞上表达(图1b)。病毒刺突与ACE2的相互作用可以被靶向刺突受体结合域(RBD)的抗体阻断,抑制病毒感染。通过与白细胞上的补体蛋白C1q或FcγR结合的抗体也可以对抗病毒感染(图1c),导致病毒和/或受感染宿主细胞直接裂解;抗体还可以促进或诱导吞噬作用,或导致有毒化学物质的释放,如细胞因子或活性氧。极少数情况下,抗体与病毒粒子的次优结合可通过抗体依赖性增强(ADE)过程促进病毒发病,其中FcγR识别病毒-抗体复合物可利于病毒进入宿主免疫细胞。使用动物抗原血清是第一种被动免疫法,但有可能引发人类抗体免疫反应,因此可使用人的血液作为抗体来源,这就是恢复期血浆疗法(CPT)。但个体之间不同的抗体反应限制了其发展,且CPT还存在安全风险,包括过敏反应和感染其他病毒(如HIV、乙型肝炎和丙型肝炎)的低风险。总体而言,CPT作用可能仅限于治疗选择有限的早期阶段。鉴于CPT的局限性,研究人员越来越关注使用中和mAb对传染病进行被动免疫。3开发人类治疗性抗体治疗病毒感染的策略人类mAb治疗病原体的策略可分为靶向已知抗原和靶向未知抗原(图2)。靶向已知抗原的mAb从噬菌体展示文库中筛选,该文库由免疫或感染个体的免疫球蛋白可变基因构建。尽管这些方法已分离出针对多种病原体(例如HIV29、SARS-CoV-2和炭疽毒素)的中和抗体,但筛选出的mAb并不代表天然抗体库。随后开发的第二种靶向方法,是直接分离抗原特异性记忆B细胞,然后识别它们产生的mAb。记忆B细胞可以来自恢复期患者的血浆,或来自携带人类免疫球蛋白基因座的转基因小鼠。图2:抗体的发现方法4抗体工程对mAb进行工程改造以改善其治疗特性被称为是抗体工程,研究表明Fc区域突变会增加或降低ADCC和/或CDC有效性,减少IgG分解代谢可改善mAb体内半衰期。工程改造的Fc区域以延长其半衰期的多种抗病毒mAb已进入临床阶段,最先进的是sotrovimab(也称为VIR-7831和GSK4182136),使用LS修饰Fc区域氨基酸,由Vir Biotechnology和GSK开发,针对SARS-CoV-2病毒,于2021年获批。Vir Biotechnology还将LS修饰用于VIR-3434,该mAb旨在预防10种乙型肝炎病毒基因型对肝细胞的慢性感染,目前处于II期试验(NCT04856085)。其他使用LS修饰的抗病毒mAb,包括用于治疗HIV的VRC01LS、VRC07-523LS和elipovimab(GS-9722)。VRC01LS是一种由美国国立卫生研究院(NIH)开发的广泛中和mAb,其血清半衰期比未改造的长约四倍,且在血清中显示相似中和活性,目前处于I期试验(NCT02256631)中。抗体工程还可用于生成新的抗体形式,例如设计用于识别两种不同表位或抗原的双特异性抗体(bsAb)。因此,单个bsAb可以与两种不同蛋白质或同一蛋白质上的两个不同位点结合。目前科学家已开发了广泛的bsAb抗体,特别是应用于肿瘤学,以及传染病研究。其他可用于抗病毒药物的新型工程抗体形式包括纳米抗体(15kDa),与小鼠和人类抗体结合域(50kDa)相比,仍保留完整抗原特异性。此外,纳米抗体能够合常规抗体通无法触及的表位,如被聚糖掩盖的保守病毒结构域。最后,锚蛋白重复蛋白(DARPins)等抗体模拟物可以提供高亲和力并和多特异性。例如,由Molecular Partners和诺华开发的一种多DARPin(ensovibep)能同时与SARS-CoV-2刺突RBD的三个单元结合,目前正在临床试验中(NCT04828161)。2抗病毒感染的抗体1抗冠状病毒,包括SARS-CoV-2冠状病毒是冠状病毒科有包膜的正义单链RNA病毒,可以感染多种哺乳动物和鸟类,导致呼吸道和/或肠道疾病。人类冠状病毒是引起普通感冒的主要原因,导致人类约30%的轻度呼吸道感染和非典型肺炎。4种不同的冠状病毒通常在人群中传播,包括HCoV-OC43、HCoV-HKU1、HCoV-NL63和HCoV-229E。在过去的二十年中,出现了3种可能导致人类生命危险疾病的冠状病毒。严重急性呼吸综合征冠状病毒(SARS-CoV)于2002年在中国出现并传播,在37个国家造成8100人感染和近800人死亡。十年后,中东呼吸综合征冠状病毒(MERS-CoV)在阿拉伯半岛出现,并传播到21个国家,感染约2500人,致死率为35%。2019年,在中国发现了一种现在称为SARS-CoV-2的冠状病毒感染,这种冠状病毒可导致发烧、严重的呼吸道疾病、肺炎、腹泻、呼吸困难和多器官衰竭。根据世界卫生组织(WHO)的数据,截至2022年2月,已确认超过4.14亿例病例,导致至少580万人死亡。所有冠状病毒都使用三聚体刺突(S)跨膜糖蛋白进入宿主细胞。S蛋白是一种I型膜I类融合蛋白,其有两个功能亚基,它们在蛋白质融合前构象中保持非共价结合(图3a)。氨基末端S1亚基负责与宿主细胞受体结合,而羧基末端S2亚基负责融合病毒和细胞膜。S1亚基进一步分为N端结构域(NTD)和RBD(图3a)。SARS-CoV-2的基因组序列与SARS-CoV有79.6%的同一性,并且与SARS-CoV相似,依靠S1亚基RBD与宿主细胞(如肺细胞和肠细胞)上的ACE2受体结合。在与宿主细胞结合后,S2亚基构象变化导致病毒融合并进入靶细胞(图3b)。S糖蛋白一直是开发靶向SARS-CoV-2mAb的焦点蛋白,它是针对SARS-CoV和MERS-CoV的有效中和mAb靶标。超过20种靶向S糖蛋白的mAb已进入临床试验,其中一些已获得FDA批准(表2)。然而,迄今为止针对COVID-19治疗的大多数mAb会因为SARS-CoV-2变体(VOC)的迅速出现而治疗受限。诸如Alpha、Beta、Gamma和Delta等VOC在S糖蛋白区域有9-12个突变,这些突变会影响mAb的治疗性。Omicron变体在S糖蛋白中累积超过35个突变,其中15个发生在RBD中,RBD不仅是与宿主受体ACE2结合的位点,也是治疗性mAb的关键靶点。Omicron VOCs(BA.1、BA.1.1和BA.2)的出现使许多mAb失效,包括casirivimab和imdevimab的联合使用、bamlanivimab和etesevimab的联合使用以及sotrovimab(表2)。不过,最近开发的两种mAb,bebtelovimab和P2G3对OmicronVOC依然能保持活性。S蛋白上的NTD结构域也是一个重要靶点(图3a),NTD特异性抗体的结构表征表明mAb靶向NTD突出区域的上侧。41个NTD特异性mAb的表位作图鉴定了6个抗原位点,其中一个被所有已知NTD特异性中和抗体识别,并被称为“NTD超位点”,由残基14-20、140-158和245–264组成,目前NTD特异性抗体起作用的中和机制仍有待确定。确定使用mAb治疗的目标人群和最佳时机至关重要。mAb被认为在具有严重COVID-19高风险的个体中发挥预防作用,如老年人和/或患有多种疾病的个体,以及对疫苗接种反应不佳或免疫功能低下的个体。最后,尽管最初担心抗SARS-CoV-2mAb会引起ADE风险,但目前没有证据表明在临床试验中任何mAb都会发生ADE。图3:针对受体结合域的SARS-CoV-2刺突蛋白和中和抗体2抗人呼吸道合胞病毒RSV是一种有包膜的负链RNA病毒,属于肺病毒科。RSV感染极为常见,通常会导致轻微的呼吸道症状。1990年代末至2000年代初,第一个名为RespiGam的RSV静脉注射免疫球蛋白制剂被用于预防患有支气管肺发育不良和早产幼儿的严重RSV相关下呼吸道疾病。RespiGam于2003年停止使用,取而代之的是帕利珠单抗预防,帕利珠单抗也是第一个用于治疗高危婴儿严重RSV感染的中和mAb,于1998年获批。在三种RSV表面蛋白(F、G和SH)中,F特异性抗体在受感染的人类血清中具有中和活性,因此F糖蛋白一直是RSV单克隆抗体开发的重点。F糖蛋白是负责将病毒膜与细胞膜融合的三聚体I型融合糖蛋白,与许多其他病毒融合糖蛋白类似。帕利珠单抗是一种人源化mAb,可与F糖蛋白的抗原位点II结合(图4a)。Motavizumab是一种亲和力高的帕利珠单抗衍生物,效力高10倍,与帕利珠单抗相比,其RSV住院率相对降低了26%。然而,2010年III期临床试验的结果显示,与帕利珠单抗相比,Motavizumab不良事件增加,导致其开发终止。用于治疗RSV的新型mAb,例如nirsevimab(MEDI8897)和suptavumab(REGN2222),Nirsevimab衍生自一种名为D25的mAb,由AIMM Therapeutics分离,靶向抗原位点Ø,在体外比帕利珠单抗更有效。3抗埃博拉病毒埃博拉病毒是属于丝状病毒科的单链RNA病毒。埃博拉病毒属包含6种不同名称:本迪布焦埃博拉病毒(Bundibugyovirus)、雷斯顿埃博拉病毒(Restonvirus)、苏丹埃博拉病毒(Sudanvirus)、塔伊森林埃博拉病毒(TaïForestvirus)、邦巴里埃博拉病毒(Bombalivirus)和扎伊尔埃博拉病毒(埃博拉病毒,EBOV)。EBOV会导致严重疾病,病死率高达25-90%。EBOV的糖蛋白是三聚体I类融合蛋白,由三个二硫键连接的GP1-GP2异二聚体在病毒表面形成三聚体。GP1亚基与EBOV受体NPC1结合,允许GP2介导的病毒和宿主细胞膜融合(图4b)。GP1带有RBS、聚糖帽和粘蛋白样结构域。GP2包含一个N端肽、内部融合环和跨膜结构域。1995年从埃博拉病毒幸存者中分离出两种mAb(mAb100和mAb114),REGN-EB3(以Inmazeb销售,再生元)是三种针对EBOV糖蛋白的完全人源mAb(REGN3470、REGN3479和REGN3471)的组合。REGN3479(现称为maftivimab)识别GP2融合环并提供中和活性,REGN3471(现称为odesivimab)识别外部聚糖帽并具有细胞靶向功能。REGN3470(现在称为atoltivimab)与GP1头部结合,提供中和活性和细胞靶向活性。图4:与来自各种病原体的糖蛋白结合的中和抗体4抗人类巨细胞病毒人类巨细胞病毒(HCMV)是一种有包膜的双链DNA病毒,其基因组大小超过235kb,是已知最大的人类疱疹病毒。HCMV是betaherpesvirus家族成员,可以在健康个体中建立终生潜伏期。原发感染一般无症状;然而免疫功能低下宿主中的病毒再激活可能危及生命,并且怀孕期间的垂直病毒传播是先天性出生缺陷的主要原因之一。HCMV利用不同的糖蛋白复合物允许细胞进入多种细胞。两种含gH / gL的复合物可调节病毒嗜性,gH / gL / gO三聚体与PDGFRα结合,是成纤维细胞感染所必需,gH / gL / UL128 / UL130 / UL131A五聚体与神经纤维蛋白2结合,是病毒进入上皮细胞、内皮细胞和骨髓细胞所必需的。(图4c)。几种靶向gH / gL或gB复合物的mAb已被分离出来,并在体外感染模型中显示出适度的功效。为了鉴定最有效的HCMV中和抗体,Lanzavechia小组从自然感染供体的记忆B细胞中分离出一大组mAb,发现五聚体复合体是HCMV中和的主要靶点,其产生的中和抗体的效力比其他HCMV复合体的效力大几个数量级。这一发现揭示了抗原选择对鉴定有效的中和mAb和设计有效疫苗的重要性。在同种异体造血干细胞移植或实体器官移植后,使用HCMV特异性mAb预防HCMV感染和疾病已得到广泛研究。MSL-109是一种靶向HCMV表面糖蛋白gH的mAb,且作为AIDS患者的HCMV诱导性视网膜炎的补充治疗,但由于缺乏疗效,在II/III期停止开发。RG7667是结合gH / gL和五聚体复合物的两种mAb的混合物,可以有效地中和所有测试细胞类型的HCMV感染。然而,在对肾移植受者进行的II期试验评估时,发现缺乏疗效。CSJ148由两种抗HCMV mAb组成,一种抗gB mAb(LJP538)和一种抗五聚体 mAb(LJP539),用于预防造血干细胞移植患者的HCMV,也终止于II期试验。因此,尽管抗HCMV mAb已做出了重大开发努力,但尚未获得FDA批准。临床上唯一可用的基于抗体的疗法仍然是Cytogam,一种用于静脉注射的CMV免疫球蛋白制剂。5抗流感病毒流感是正粘病毒科的一员,有4种类型——甲型、乙型、丙型和丁型流感——它们都有一个分段的负义单链RNA基因组。甲型和乙型流感是造成人类严重感染的原因,而丙型流感仅引起轻微症状,丁型流感不会感染人类。该病毒基因组由编码至少12种蛋白质的8个片段组成:血凝素(HA)、神经氨酸酶(NA)、PB2、PB1、PB1-F2、PA、PA-X、NP、M1、M2、NS1和NS2。病毒表面的两种糖蛋白是HA和NA,HA比NA更丰富(每个病毒粒子约500个分子),比例为4:1至5:1。HA头部是发生抗体反应的主要目标,其序列和糖基化位点变化多样。尽管HA的结构总体上是保守的,但其序列和糖基化模式在不同亚型之间不同。由于其高序列可变性,针对免疫优势头部的抗体反应是菌株特异性的,并且仅提供短期免疫。其头部含有RBS,它通过与唾液酸受体结合将病毒附着到宿主细胞上。RBS形成一个浅口袋,由4个部分组成:190螺旋和130、150和220环,除了220环外,RBS本身相对保守,而其余的头部在序列上是高度多样化的。抗体通常针对5个主要抗原位点,即H1的Ca1、Ca2、Cb、Sa和Sb,以及H3的位点A-E,它们位于RBS周围。RBS位点的抗体可以阻断病毒附着或阻止受体介导的内吞作用,因此中和病毒。HA茎部主要在亚型内高度保守,并且在某种程度上跨亚型,以比头部结构域慢得多的速度进化(图4d)。MEDI8852是一种人IgG1 mAb,从一名无并发症的甲型流感感染患者中分离出来,目前处于IIa期。Vir Biotechnology正在开发VIR-2482,这是一种中和甲型流感的mAb,它与HA的保守区域结合并能中和自1918年西班牙流感(H1N1)以来的所有主要毒株。因此,这种类型的抗体可用作通用预防剂,克服当前流感疫苗的局限性。此外,由于VIR-2482已通过改造其Fc区域延长血清半衰期。3对疫苗开发的影响1用于疫苗设计的抗原-抗体相互作用和结构疫苗学疫苗已被证明是传染病最有效的预防策略。然而,依赖三类传统疫苗(减毒活疫苗、灭活疫苗和解离病原体)对HIV、RSV、流感、丙型肝炎病毒、HCMV或EBOV等病毒已不起作用。对于大多数疫苗而言,抗体反应至关重要。因此,鉴定能够有效中和病原体的抗体是加速疫苗开发的关键因素。反向疫苗学2.0,也称为基于抗体的疫苗学,旨在克服传统方法的局限性,通过基于抗原与其同源抗体复合的结构表征设计新型疫苗(图5)。通常通过对感染细胞的粗匀浆进行蛋白质组学分析来鉴定抗原,然后根据免疫原性及其刺激免疫反应的能力进行选择。然而,这种方法非常耗时,并且在鉴定新的蛋白质时不能保证其结构完整性或免疫原性。实验驱动的中和mAb的分离和其靶标鉴定,仍然是鉴定候选疫苗的最佳方法。抗体驱动和基于结构的抗原设计策略在开发新出现的传染病疫苗方面特别有效。图5:反向疫苗学2.02人呼吸道合胞病毒对于RSV,中和抗体主要针对融合(F)蛋白的头部结构域(图4a),为了集中针对F头部抗原位点的免疫反应,Boyington等人设计了仅包含头部区域的截断F免疫原。这些蛋白质在融合前状态下特别脆弱,因此需要稳定其结构,将抗原锁定在融合前构象中,以促进阻止膜融合的中和抗体反应。这种结构信息可用于识别稳定突变,多种基于结构的设计策略已被应用来稳定F蛋白的融合前构象。基于结构的疫苗设计中一个相对年轻的领域是表位支架,它依赖于将病毒表位移植到不相关的载体蛋白上,即所谓的支架蛋白,以将免疫反应集中于保守的、功能性的位点上,这些位点是中和抗体的靶标。这一策略首先应用于设计新的HIV免疫原,由McLellan等人首次实现,他们通过将莫他珠单抗的表位移植到不相关的蛋白质支架上,将F蛋白的抗原位点II设计成免疫原。3埃博拉病毒多个平台的疫苗已显示出对EVD的保护作用,其中两种并非基于反向疫苗学概念的疫苗已获得许可:包括Ervebo(rVSV-ZEBOV)和Zabdeno / Mvabea的联合使用。这两种疫苗都基于病毒载体,最近对接种疫苗的人类B细胞反应的分析表明,疫苗诱导的mAb中的体细胞超突变水平较低。基于蛋白质的亚单位疫苗的合理设计工作正在进行中。其他正在开发的疫苗包括cAd3-EBOZ,一种黑猩猩腺病毒(cAd3)的减毒型,编码EBOV-GP糖蛋白。最后,发现含有EBOV-GPMakona株的重组纳米颗粒疫苗可诱导有效的免疫反应。4人类巨细胞病毒HCMV疫苗的开发始于1970年代初期,实验室工作分离了两种减毒病毒株:AD169和Towne。AD169减毒株很快被放弃,Towne减毒株在实体器官移植受者和健康志愿者中进行了广泛测试。寻找CMV疫苗的关键是鉴定CMV的表面蛋白,称为糖蛋白B或gB(图4c)。当与MF59水包油佐剂联合使用时,该疫苗是安全且部分有效的,但在人体中3次注射后中和抗体水平较弱。亚基gB蛋白还与AS01佐剂结合以刺激TLR4受体,从而在人体中引发更高和更长时间的抗gB抗体水平。gB抗原是III类三聚体融合蛋白并且在融合后构象发挥作用,有人提出融合前构象中的gB可以产生更高的中和反应,但最近的结果表明可能并非如此。CMV表面存在的蛋白质五聚体复合物由糖蛋白H(gH)、糖蛋白L(gL)和基因UL128、130和131A的产物组成,该复合物已被证明能产生比gB更高滴度的中和抗体(图4c),这一发现推动了HCMV疫苗领域的发展。然而,如果需要中和抗体来预防HCMV的感染和传播,则还需要强烈的T细胞反应来抑制血清阳性患者的病毒再激活。即将进行的使用重组蛋白亚基(NCT05089630)或mRNA(NCT05105048)的临床试验将表明五聚体是否可以单独使用,或者是否需要与gB一起使用。5流感病毒开发流感疫苗的主要障碍之一,是病毒附着的高度变化的HA头部的免疫优势。因此,基于结构的设计方法一直专注于HA茎区(图4d)。为了将抗体反应转移到HA茎结构域,对嵌合HA分子进行了改造,该分子由与不同HA头配对的共同HA干组成。小鼠的反复免疫表明可以产生HA干特异性抗体反应,从而产生异源和异源亚型免疫。在雪貂中,这些构建体显示出在流感病毒攻击后减少病毒载量。嵌合HA已在I期临床试验中进行测试,证明其安全性,以及能够诱导广泛、强烈、持久和功能性的免疫反应。也有设计策略通过去除HA头产生分离的HA干抗原,如Yassine等人的无头HA和Impagliazzo等人的mini-HA设计。6冠状病毒获取SARS-CoV-2 S蛋白的结构数据在开发多种COVID-19疫苗(包括非常成功的mRNA疫苗)中发挥重要作用。开发有效疫苗的另一个关键方面,是从HCoV-HKU1和MERS-CoV等其他冠状病毒的S蛋白研究中获得丰富经验。7艾滋病病毒HIV疫苗开发研究极大受益于人类bNAb的分离,使用bNAb指导的HIVEnv三聚体稳定化是许多免疫原的基础。此外,最有希望的策略是使用稳定的三聚体,来激活特定的产生bNAb细胞前体并引导它们产生成熟的bNAb——这种策略称为种系靶向。最近,在接种eOD-GT8免疫原的健康受试者中报告了97%的血清转化,展示了种系靶向策略的强大潜力。4总结与展望SARS-CoV-2的出现强调了传染病的严重性,短时间内几种抗COVID-19疫苗和mAb疗法的发现和开发、制造和监管评估方面取得了前所未有的成功。这些进步是基于过去几十年在免疫学和疫苗学方面取得的巨大科学进步,也是基于对冠状病毒生物学重要知识的了解。利用抗体来治疗COVID-19也受益于技术进步和专业知识完善,但mAb有一定局限性,如SARS-CoV-2 Omicron VOC会导致抗体的病毒靶标发生突变以致病毒逃脱中和。希望本文和其他地方总结的近期临床成功药物和案例研究,对抗感染mAb的研发能有所启示。鉴于目前mAb的一个关键限制仍然是其在组织中的有限分布,将mAb特异性递送至靶标的新型的递送策略,可能会使mAb成为对抗病毒感染的更强大工具。
抗体疫苗免疫疗法信使RNA
2022-07-30
摘要:在以靶点优先的抗体生成方法中,需要对靶点特质、试剂质量、宿主选择等各方面因素进行综合考虑。同时也有以噬菌体展示技术为代表的各种抗体分离技术,本文详述了靶点选择中需要考虑的各个因素以及各种抗体分离方法的优缺点,继续讨论为生产ADC药物研发理想抗体的高效策略。靶点在抗体筛选中的考虑因素(1)高质量试剂高质量试剂对抗体生产至关重要。许多用于靶点验证的试剂(如靶DNA、表达质粒、天然和重组细胞系、纯化蛋白和对照或参考抗体)在抗体生成和筛查过程中也同样可能有用。当生产筛选试剂时,平行生成可用于检测抗体hits物种交叉反应的靶同源基因至关重要。(2)靶点性质选择用于ADC干预的靶点性质和固有特性大相径庭。在肿瘤细胞和肿瘤血管中,几乎所有的基因都能在细胞表面表达。它们是完整的膜蛋白,通常含有一个或多个跨膜α螺旋,并会永久附着在细胞膜上。这种蛋白质只能用洗涤剂、非极性溶剂或变性剂从生物膜上分离。出于这个原因,绝大多数用于抗体产生和筛选的蛋白质抗原会被分解成较小的可溶性亚区,或细胞表面受体的胞外区。这些片段可以作为免疫原或选择试剂进行重组表达设计。精确确定结构域边界通常至关重要,因为表达结构中任何微小变化都可能对表达量产生巨大影响。(3)宿主选择可以用原核或真核宿主重组生产目的抗原。宿主选择可能取决于靶点性质。最重要的是,产物应该是均质、适当折叠的抗原,以便获得关键表位。为了快速生成,最好在HEK293或CHO细胞中瞬时表达。许多蛋白质标签(如流感血凝素(HA)、FLAG或His6),能从复杂的细胞培养液中轻松纯化。因为Fc结构域可以独立折叠,在体内外都能改善配对分子的溶解性和稳定性,同时也为蛋白A/G亲和层析提供了一个方便的纯化手柄。所以也可以使用由基因上融合到靶抗原的免疫球蛋白Fc结构域组成的基于Fc的融合蛋白。(4)考虑使用过表达细胞系进行筛选表达肿瘤学靶点的转化细胞系通常易于生产。这些细胞系是抗体生产的有效免疫原,在动物中能产生强烈的免疫反应。整个细胞都是由许多细胞表面蛋白组成的复杂抗原,每个蛋白都是一种潜在的抗原。因此,需要通过广泛筛选来生产靶向单一抗原的特异性抗体。在基于细胞的筛选过程中,为了降低复杂性,可以使用重组过表达细胞系。全长靶抗原可以在常用的细胞系统中表达,如HEK293或CHO细胞。非转基因、低表达或不表达靶点的亲本对照细胞可用于反筛选。(5)小鼠细胞系表面过表达为了最大化增强小鼠在全细胞免疫时的特异性抗体反应,也可以在来自与免疫宿主同源的菌株的小鼠细胞系的表面过表达人类靶抗原。与人类细胞不同,表达重组抗原的小鼠细胞背景反应降低,因为它们的整个细胞表面成分(除转染靶分子和表达靶分子外)都源于小鼠,因此在动物体内是耐受的。可能使用自然表达的肿瘤细胞系和重组蛋白,以及不同背景下的工程细胞系来生产理想抗体。抗体分离的几种方法(1)杂交瘤技术目前市场上的许多治疗性抗体是使用Kohler和Milstein开发的传统杂交瘤技术生成。杂交瘤细胞系是由永生化的骨髓瘤细胞与产生免疫球蛋白的B细胞稳定融合而产生的,这本身效率不高。尽管存在局限性,杂交瘤在历史上一直是最广泛使用的抗体生成方法。但最近,已采用能更有效识别抗体的方法,包括使用CD40/CD40L培养和扩增免疫啮齿动物产生抗体的B细胞。以及基于流式细胞术的抗体产生细胞(APC)的分类,使用荧光标记的抗原,然后从细胞中进行逆转录聚合酶链式反应(RT-PCR)。(2)转基因技术上述每一种方法都依赖于对啮齿动物或其他动物的免疫,并将此作为产生抗体的免疫细胞的来源。小鼠由于其世代周期短,遗传标记(genetic markers)丰富和饲养管理成本低,经常被用于免疫。使用靶向缺失特定基因的基因敲除小鼠,可以更容易地提高对某些蛋白质靶标的免疫反应,这些蛋白质靶点在物种间高度保守。当没有基因敲除小鼠时,可以使用大鼠或其他物种来促进人/鼠交叉反应抗体的产生。转基因啮齿动物可以在不需要人源化的情况下,相对快速地生成靶向已验证靶点的抗体。但转基因技术成本较高和转基因动物数量有限,所以大多数研究人员更偏好非转基因动物。(3)DNA免疫图1 尾静脉注射流体动力法DNA免疫及其免疫应答特征(来源于:Hazen et al 2014))最近对该技术的改进表明,DNA免疫能够产生靶向G蛋白偶联受体(GPCRs)和离子通道等抗原的免疫反应,这些抗原通常更难产生抗体。一般来说,DNA免疫所产生的免疫反应比其他方法所产生的免疫反应需要更长的时间,但对于更传统的方法来说,延迟的反应时间通常能抵消产生免疫原所需时间。(4)快速免疫方法--RIMMS能快速生成一组针对多个靶点的抗体对于ADC靶点验证非常有效。对啮齿动物免疫通常需要两个月或更长时间,才能获得足够的抗体效价。但利用快速免疫方法,如重复免疫多个部位(RIMMS), RIMMS即多位点重复免疫(repetitive immunizations multiple sites)意思是在相同的地方低剂量短间隔重复免疫,一般是在小鼠背上皮下选择四五个点,每三天左右在这些点进行低剂量免疫,一般十几天就可以完成免疫。这种方法可以在不影响质量的情况下,缩短免疫时间。RIMMS主要依靠次级淋巴组织中早期发生的快速超突变和亲和力成熟来对抗抗原。2-3周内,分别在几个皮下部位注射低浓度抗原,每周三次。在抗原攻击后,收集每个注射部位的引流淋巴结,并用于融合或基于B细胞的抗体筛选。(5)噬菌体展示技术噬菌体展示技术为将抗体引导到具有特定特征的抗体群体,例如,靶向结合到一种肿瘤细胞,靶向与已知靶点上的已知表位结合的抗体、或二者兼具。噬菌体来源的抗体通常比免疫动物来源的抗体亲和力低。但由于上述结合部位屏障的考虑,以及中等亲和力抗体可能会避免被内化到低表达的正常细胞,虽然仍可能在高表达的肿瘤细胞中内化,但纳米摩尔数较低抗体很可能适用于ADC。图2 亲和力与抗原内化率有关,决定了抗体在体外分解代谢的程度。将450 ng~(125)I标记的抗HER2抗体与5×105个SK-OV-3细胞孵育,进行内化试验。在未结合的免疫球蛋白从细胞中清洗后的1、2、4、8、24小时后立即采集样本相对于表面结合的时间,表示总抗体的百分比(A),解离(B),内化(C),抗体降解(D)三份样品的平均标准偏差为±1。降解与抗原内化的相关性(E)。灰色区域表示由其他人确定的内化率(Ke)范围1固液选择使用纯化或重组抗原在溶液或固相中进行噬菌体选择。随后筛选与表达在工程细胞或肿瘤细胞表面的抗原结合的抗体。这种方法的优点在于抗体靶点特异性是已知的,而与更复杂的细胞表面抗原结合只是用于确认。此方法主要的缺点在于需要产生高质量的蛋白质试剂。即使在成功选择之后,被选择与纯化抗原结合的抗体经常无法识别肿瘤细胞背景下的靶点。选择与细胞结合的噬菌体,包括肿瘤细胞系、肿瘤组织样本或过表达特定靶点的哺乳动物表达系可降低克隆与细胞表面靶标结合失败的风险,但随着细胞表面复杂性的增加,分离与靶点结合的克隆与不与靶点结合克隆难度也会逐渐提高。图3 固液筛选原理 2细胞筛选对过表达蛋白进行细胞选择也是分离已知特异性抗体的可行途径。这种方法有利于分离与已知靶点的细胞表面形式特异结合的抗体。建立稳定的哺乳动物细胞系可能非常耗时,但通常转基因的细胞系及其未转基因的亲本可用于抗体的选择和筛选,并为靶标特异性提供明确的证据。重组细胞系可能高估了低亲和力抗体与表达较低靶标的细胞结合的能力,并可能改变了内化机制、糖基化或与其他蛋白质的结合,因此验证肿瘤细胞上的抗体属性是必要的。图3 不同筛选体系中表位暴露示意图通常,可以结合各种方法优势。如果靶点已知,在纯化的蛋白质和细胞上交替进行噬菌体筛选可以提高所产生的克隆与肿瘤细胞上呈现的所需靶点特异结合的可能性。同样利用对纯化蛋白的选择和对肿瘤细胞的内化选择,可以将选择的池集中在所需表型的克隆上。很难预测展示在噬菌体上的Fab单链抗体的哪些特征将转化为免疫球蛋白G(IgG)形式的特征。基于这个原因,通常推荐分离物转换成免疫球蛋白来确认它们的结合和内化特征。结 语在ADC药物研发过程中,抗体是核心也是灵魂。靶点优先的抗体生产方法基于对靶点的先验知识在抗体生产过程中拥有更大的独立性和新颖性,也更容易设计楚理想受体。同时以噬菌体展示技术、重复免疫和DNA免疫为代表的各种抗体分离技术也为高效获得理想抗体,提高免疫效率等进行了有益尝试。
抗体抗体药物偶联物免疫疗法
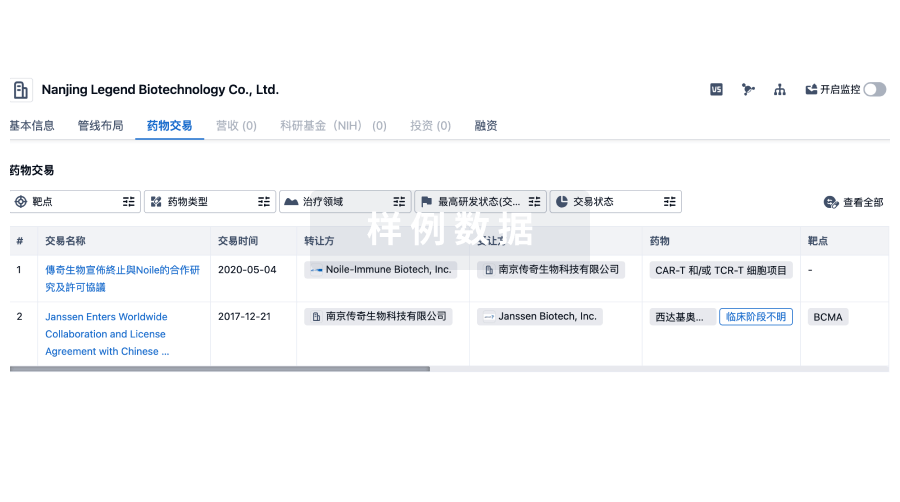
100 项与 Milstein Dermatology LLC 相关的药物交易
登录后查看更多信息
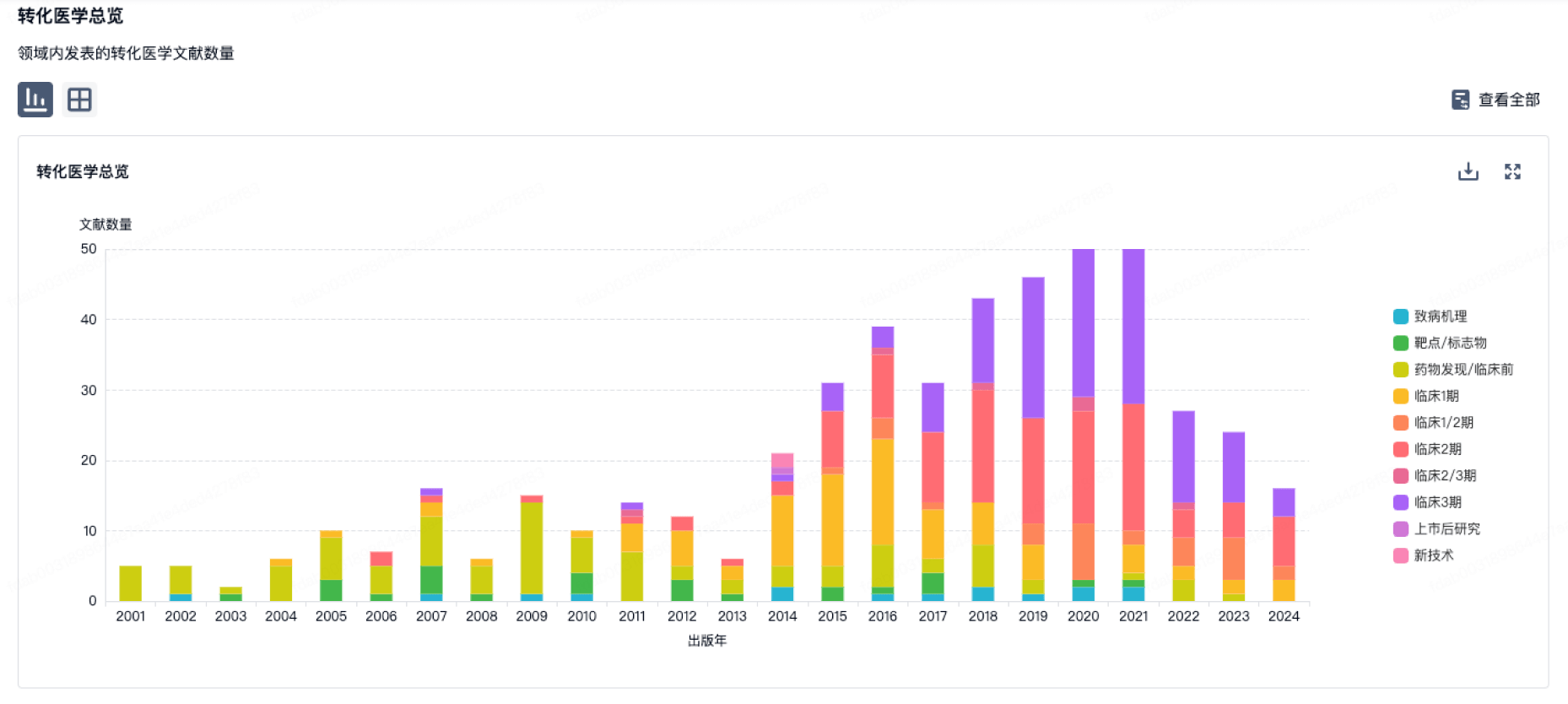
100 项与 Milstein Dermatology LLC 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2024年07月08日管线快照
无数据报导
登录后保持更新
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
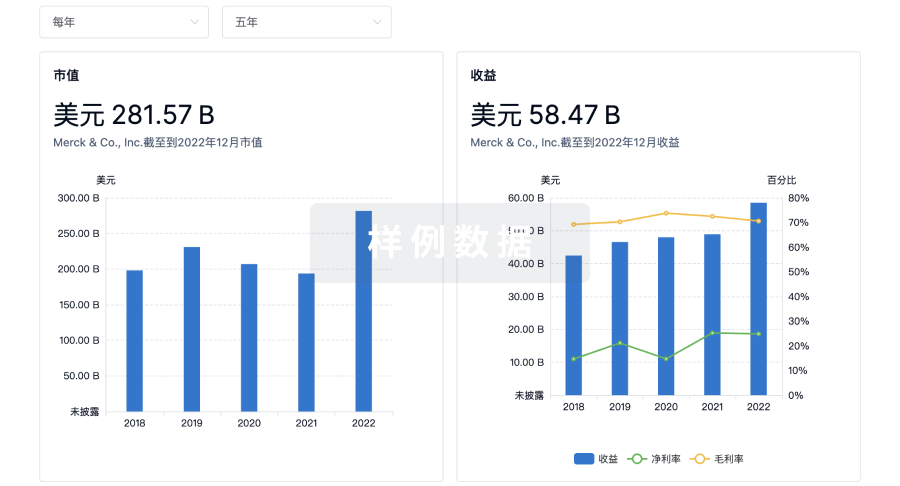
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
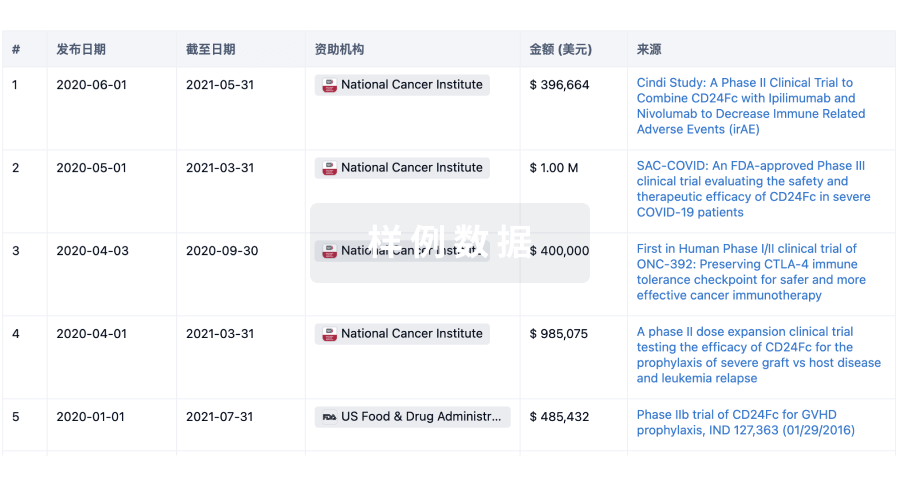
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
标准版
¥16800
元/账号/年
新药情报库 | 省钱又好用!
立即使用
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用