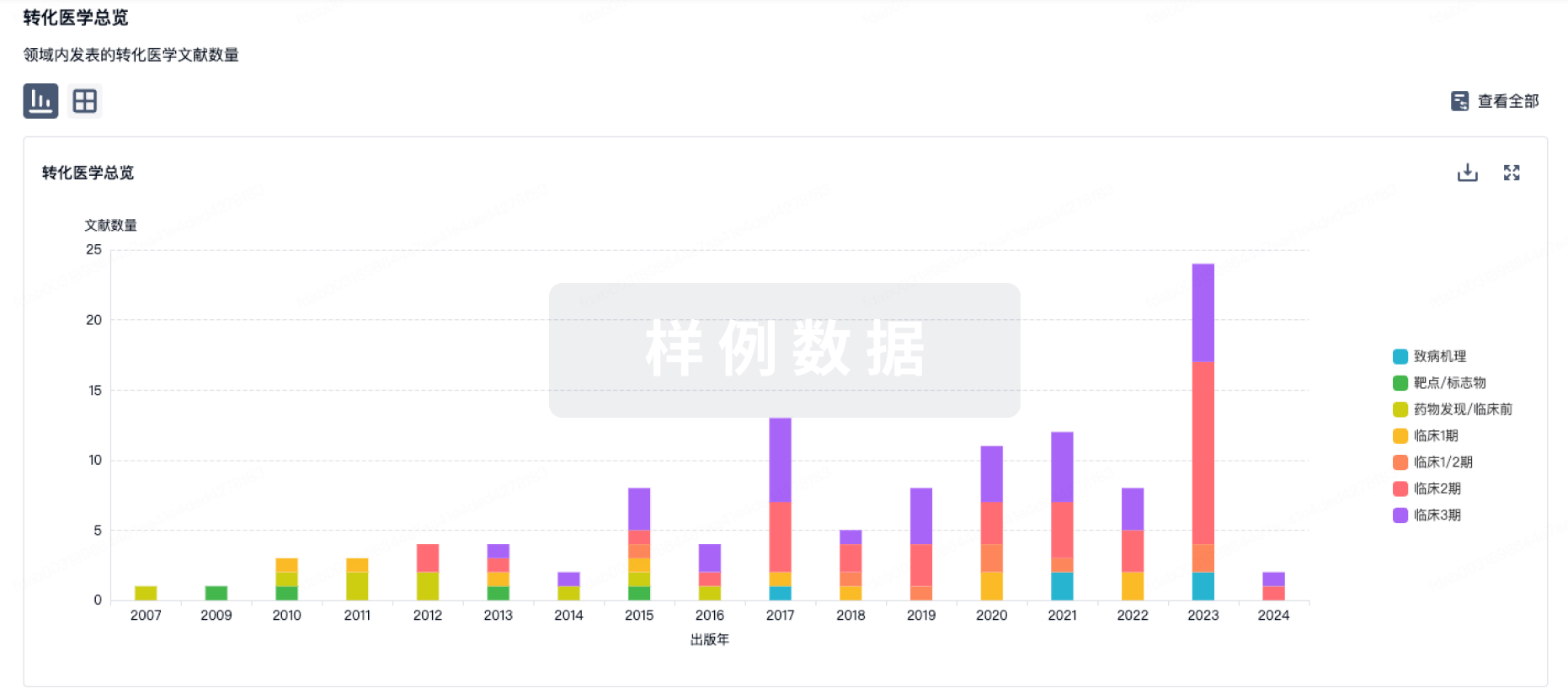
“不可成药”蛋白质通常是一类具有大而复杂的结构或功能的蛋白质,难以使用常规药物设计策略干扰其表达。靶向这类蛋白目前被认为是治疗人类疾病的潜在机会,在医学领域吸引了不少关注。2023年9月发表在Nature子刊《Signal Transduction and Targeted Therapy》上的最新综述,全面介绍了针对“不可成药”蛋白的药物发现的最新进展及其在临床上的应用。近些年,随着基因组学和蛋白组学的兴起,在人类疾病中发现了许多具有临床意义的靶点。然而,传统的“可成药”靶点占比较少,许多靶点由于功能界面平坦且缺乏用于配体相互作用的明确口袋,被归于“不可成药”靶点,这使得药物设计面临巨大挑战。随着对“不可成药”靶点的研究不断深入,具有相似不可成药特征的各种分子被分为以下几类:1.小GTP酶RAS家族蛋白,包括KRAS,HRAS和NRAS,属于小GTP酶。这些蛋白由于表面缺乏可靶向的口袋,被认为是不可成药的。2.磷酸酶根据结构特征,磷酸酶被分为两种类型:蛋白质酪氨酸磷酸酶(PTPs)和蛋白质丝氨酸/苏氨酸磷酸酶(PSTPs)。由于每种磷酸酶的结构存在很多相似性,具有选择性低和副作用难以避免的问题,极大地阻碍了药物发现的进展。3.转录因子(TFs)多种人类疾病与参与许多生物过程的TFs失调有关,其中大多数由于其结构异质性和可处理结合位点的缺乏而无法被常规小分子靶向。值得关注的TFs包括参与肿瘤病理过程的p53,Myc,ER,AR,XBP1,年龄相关疾病和神经退行性疾病中的NRF2,以及免疫疾病中的NF-κB,BTB, BACH,EB,E3目前的研究主要集中在靶向p53和Myc上。4.表观遗传靶点表观遗传靶点在调节基因表达模式中起着至关重要的作用,并在各种生物过程和疾病中产生影响。表观遗传修饰的主要类型包括DNA甲基化,组蛋白修饰,非编码RNA,染色质重塑和其他表观遗传酶。靶向这些表观遗传靶点有可能揭示各种疾病的潜在机制。5.其他蛋白质蛋白质-蛋白质相互作用(PPI)及其网络在生物过程和细胞周期调节中具有重要意义,为治疗复杂疾病提供了另一种潜在途径。RAS和TFs如p53和Myc也受到PPI网络的影响。一部分具有平坦相互作用表面的PPI比其他PPI更难靶向,使它们在一定程度上“不可成药”。经典的PPI相关蛋白Bcl-2家族的抗凋亡成员。此外,由于缺乏结合腔,具有高度动态结构的固有无序蛋白质与各种蛋白质伴侣相互作用,也被认为是不可成药的PPI蛋白。如今,面对所谓的“不可成药”靶点,学术界开发了数十种创新方法,根据不可成药蛋白的机理,采用基于片段的药物发现(FBDD),计算机辅助药物设计(CADD),虚拟筛选(VS),DNA编码文库(DEL)等尖端技术,形成了系统的药物设计策略。01共价调节共价抑制剂,也称为不可逆抑制剂,是一类通过将轻度反应性官能团形成的共价键与靶蛋白的氨基酸残基结合以赋予额外的亲和力的抑制剂,共价结合的靶标可以被持续抑制,直到蛋白质降解和再生。与非共价抑制剂相比,共价抑制剂具有持续抑制和滞留时间长的优点。同时,共价抑制剂还可以减少用量,提高顺应性,避免一些潜在的耐药机制。例如,已获准用于COVID-19的Paxlovid中的Nirmatrelvir是SARS-CoV-2主要蛋白酶Mpro的抑制剂,突出了半胱氨酸反应性共价官能团在靶向蛋白酶活性位点中的重要性。此外,共价抑制剂可以靶向缺乏表面“口袋”的不可成药蛋白质,从而有可能扩大治疗范围。在这一领域,KRAS抑制剂Sotorasib的批准是共价药物开发进展的一个里程碑。图1 针对不可成药蛋白的共价调节剂1.共价KRAS抑制剂KRAS在参与细胞生长和存活的信号通路中起着至关重要的作用,它是RAS家族中最主要的突变亚型,特别是与胰腺癌,结直肠癌和肺癌的发生密切相关。KRAS蛋白会在失活和激活状态之间转变,当KRAS与GDP结合时处于失活状态,当它与GTP结合时处于激活状态,这一转变受两类因素的调节:(1)鸟嘌呤核苷酸交换因子(GEF),例如SOS蛋白,它们催化KRAS和GTP结合状态之间的过渡;(2)GTP酶活化蛋白(GAPs),促进与KRAS结合的GTPs的水解,从而抑制KRAS的活性。直接靶向KRAS存在许多困难,首先RAS蛋白的正常活性是维持正常细胞功能所必需的。此外,KRAS与NRAS和HRAS具有很高的同源性,如果KARS抑制剂的特异性不足,可能会抑制NRAS和HRAS的活性,大概率会产生副作用。目前已知的KRAS活性功能域(药物的潜在作用位点),主要是KRAS与GDP、GTP结合的口袋状功能域。相较于蛋白激酶与ATP的较弱亲和力,KRAS与GTP、GDP的结合性非常强,亲合系数达到皮摩尔浓度级,而人体细胞组织里的GDP/GTP浓度远高出KRAS结合所需的浓度。综上所述,KRAS蛋白是一种无特征的近球形结构,没有明显的结合位点,因此难以合成能够有效靶向和抑制其活性的化合物。KRAS中常见的突变位点包括密码子12、13和61,密码子12是最常见的突变位点。近年来,亲电优先方法(electrophile-first approaches)的突破使得靶向KRASG12C突变体成为可能。半胱氨酸位于KRASG12C的密码子上,可以选择性地靶向突变体KRAS。重要的是,KRAS活性位点缺乏半胱氨酸,KRASG12C可以以共价方式实现特异性抑制。在KRASG12C突变体中,与突变体胱氨酸共价结合的小分子更容易与GDP结合的KRAS蛋白结合。这种结合降低了GTP和KRAS之间的亲和力,从而阻止了GEF催化GTP取代GDP,从而将KRASG12C突变体锁定在失活状态。KRASG12C突变体上这种结合“口袋”的发现加速了几种专门针对KRASG12C突变体的小分子共价抑制剂的开发。其中,Sotorasib和Adagrasib已在临床使用,还有有十多种正在进行临床试验。已上市的用于抑制KRAS的共价药物Sotorasib(AMG-510)2021年5月,FDA批准靶向抗癌药物Lumakras(Sotorasib,AMG-510)用于治疗非小细胞肺癌患者的KRASG12C突变,它也成为世界上第一个治疗KRAS基因突变的靶向药物,打破了“无法成药”的困境。Sotorasib通过与GDP结合并将KRAS锁定在非活性状态,特异性且不可逆地抑制KRASG12C。此外,Sotorasib已被证明能强烈抑制KRASG12C细胞中ERK蛋白的磷酸化,从而抑制细胞增殖。目前关于Sotorasib的研究已经确定了其可用于治疗腺癌、转移性结直肠癌和转移性非小细胞肺癌等,已先后在欧盟、日本等国家获得上市批准。根据最新的ACCR报告,Sotorasib治疗KRASG12C突变NSCLC患者的总缓解率为37%,疾病控制率为81%,中位无进展生存期为6.8个月,中位缓解持续时间为10.0个月。Adagrasib (MRTX - 849) Mirati Therapeutics和Array BioPharma合作鉴定了另一种不可逆的KRASG12C小分子共价抑制剂Adagrasib (MRTX-849) 。Adagasib与KRASG12C的Cys12共价结合,并延伸到变构口袋S-II P,从而将KRAS蛋白锁定为非活性构象,并抑制RAS/MAPK激酶信号传导。Adagrasib对KRASG12C的选择性比野生型KRAS和其他含有Cys的蛋白高1000倍以上。口服生物利用度高达30%,单次服用后半衰期为25小时。目前,Adagrasib的研究已经揭示了其治疗晚期实体瘤、转移性结直肠癌、转移性NSCLC和转移性胰腺癌的潜力。Adagrasib于2019年1月进入III期临床试验,根据最新的ACCR报告,它的总缓解率为58%,中位治疗持续时间为9.5个月,中位缓解持续时间为12.6个月。2022年12月12日,FDA加速批准Adagrasib用于识别KRASG12C突变的局部晚期或转移性NSCLC成年患者。2.共价EGFR抑制剂表皮生长因子受体(Epidermal growth factor receptor, EGFR)是酪氨酸激酶受体家族的一员,是一种典型的跨膜受体,在配体刺激的二聚化过程中启动信号级联反应,从而激活其酪氨酸激酶和多种下游效应物。此外,它还参与胚胎发生和干细胞分裂,并与细胞增殖、有丝分裂和癌症的发生有关。野生型EGFR蛋白的过表达或活性升高可通过信号级联导致细胞增殖、迁移、存活和抗凋亡,与非小细胞肺癌、乳腺癌、胶质瘤、头颈癌、宫颈癌、膀胱癌等多种癌症的发生发展密切相关。因此,EGFR已成为设计和开发抗癌药物的一个有希望的靶点。EGFR的靶向药物是酪氨酸激酶抑制剂(tyrosine kinase inhibitors, TKIs),它抑制细胞质中的激酶,从而阻止它们激活EGFR信号通路。第一代TKIs,如吉非替尼和厄洛替尼,以非共价键选择性结合EGFR的ATP结合位点,从而抑制EGFR磷酸化,显著延缓非小细胞肺癌的靶向治疗疾病进展。然而,耐药现象逐渐出现:只有10-19%的晚期非小细胞癌患者对吉非替尼有肿瘤反应;在使用第一代TKIs约9-14个月后,几乎所有肿瘤再次进展。随后的研究表明,非小细胞肺癌患者对吉非替尼或厄洛替尼的敏感性降低与EGFR特异性激活突变有关。突变的EGFR限制了可逆性EGFR TKIs的疗效并使其无法成药。为了克服这个问题,不可逆的EGFR TKIs,即第二代EGFR TKIs,被设计成与结合位点共价结合,从而增强对肿瘤细胞的持久抑制。与第一代EGFR TKIs相比,第二代EGFR TKIs,如阿法替尼、达康替尼和纳拉替尼,具有丙烯酰胺Michael受体侧链,可以在ATP结合位点不可逆地与Cys797结合,在临床实践中表现出更强的抑制作用。然而,第二代EGFR抑制剂仍然不能用于治疗第一代EGFR抑制剂后发生耐药突变的患者,并且也可能导致耐药,这些耐药通常与T790M突变相关。因此,第三代EGFR TKIs如WZ 4002、奥西替尼和罗西替尼,它们是专门为T790M突变体而设计的。第三代药物保留丙烯酰胺基团,与Cys797共价结合,但用嘧啶取代了第一代和第二代化合物的喹唑啉部分,提高了对T790M的选择性,对T790M的亲和力高于WT-EGFR.因此,开发共价EGFR抑制剂是非常有吸引力的。目前市场上有几种EGFR TKIs,其中第二代TIKs和第三代TKIs为共价抑制剂。已上市的抑制EGFR的共价药物阿法替尼 (GiotritTM, BIBW-2992)是FDA批准用于肺癌的第一个共价EGFR抑制剂,属于第二代EGFR TKI,于2013年7月上市。与第一代EGFR TKI类似,阿法替尼在铰链区与Met793主链形成氢键,并与疏水区相互作用。呋喃基暴露在溶剂中,3-氯-4-氟苯基位于“守门人”残基附近。临床试验表明,对于EGFR外显子缺失的患者,阿法替尼的总生存期优于化疗,与第一代EGFR抑制剂吉非替尼相比,阿法替尼总体上提供了更长的有效治疗期和良好的疾病控制。此外,阿法替尼与HER2的Cys805共价结合,被称为泛HER2抑制剂。然而,阿法替尼通过抑制WT-EGFR表现出剂量依赖性细胞毒性,并导致对吉非替尼和厄洛替尼的耐药。此外,9.1%的EGFR突变NSCLC患者发现EGFR外显子20插入(ex20ins)对阿法替尼不敏感。达克替尼(PF-299804),也是不可逆的第二代EGFR TKI,最初由辉瑞公司开发,由SFJ制药公司于2012年共同开发,于2018年被FDA批准用于治疗转移性NSCLC的外显子19缺失和外显子21替代。达克替尼具有与阿法替尼相似的结合性质,与铰链残基形成氢键,并与结合口袋中的铰链残基形成疏水相互作用。此外,在一项随机的III期临床试验(ARCHER 1050)中,与吉非替尼相比,达克替尼的无进展生存期更优;但是,观察到了更严重的不良反应。作为EGFR突变敏感型NSCLC的一线药物,达克替尼已被许多临床试验证明可延长总生存期,并显示出优于第一代EGFR TKI的显着优势。因此,FDA于2018年9月批准达克替尼用于晚期NSCLC的一线治疗。奥希替尼(AZD9291)是目前获批临床使用的第三代不可逆EGFR TKI,于2015年11月获得FDA加速批准用于NSCLC的二线治疗,随后在2018年获得FDA批准用于一线治疗。基于嘧啶环,奥希替尼通过不饱和烯丙基链形成共价键,靶向ATP结合位点的Cys797残基,从而不可逆地结合EGFR激酶的催化活性中心,抑制EGFR及其下游信号底物Akt和Erk的磷酸化。此外,多项研究表明,奥希替尼能够有效穿透血脑屏障,对晚期NSCLC的BMS具有良好的治疗效果,并显着延长中枢神经系统转移病例的无进展生存期。除了单独使用外,奥希替尼还正在与其他NSCLC靶向治疗药物联合使用,例如Met,Bcl-2和MAPK途径的抑制剂。由于奥希替尼的显著疗效,在奥希替尼临床开发的同时,几种基于奥希替尼结构的第三代EGFR TKIs也在开发中。奥莫替尼 (HS-10296)是由Hansoh制药公司开发的口服,不可逆的第三代EGFR TKI。在奥希替尼的基础上进行结构优化,引入环丙基取代吲哚环上的甲基,该环丙基可与Met790侧链形成疏水相互作用。该优化提高了抑制活性和WT-EGFR选择性,同时增加了亲脂性和血脑屏障通透性。它于2020年3月在中国获得批准,并在2022年的III期试验(NCT03849768)中被证明是一种耐受性良好的第三代EGFR TKI,可作为EGFR突变NSCLC的一线治疗选择。然而,奥莫替尼对突变型EGFR仍然具有更强的选择性,耐药或致敏EGFR的半抑制浓度值比野生型酶低约2-16倍。Lazertinib (YH-25448)是 Genosco 开发的具有强大血脑屏障穿透力的不可逆第三代 EGFR TKI,已被证明可诱导 EGFRL858R+T790M突变小鼠皮下和颅内病变的剂量依赖性消退尽管观察到不良反应,但与相同剂量的奥希替尼相比,它在抑制肿瘤生长和改善总生存率方面具有优越的功效。2021年1月,获得韩国非小细胞肺癌治疗上市许可。阿氟替尼(Furmonertinib)是中国自主研发的专门针对EGFR突变的第三代药物,是奥希替尼的优化版本。阿氟替尼加入2,2,2-三氟乙基取代甲基,并引入N原子以吡啶环取代苯环。保留的Michael加成受体-丙烯酰胺结构允许阿氟替尼与Cys797残基共价结合,产生有效的抗肿瘤作用。此外,阿氟替尼的氨基嘧啶主环可以克服T790M突变引起的位阻,而三氟乙氧基吡啶结构的引入则阻断了非选择性代谢物的产生。这增强了阿氟替尼的活性和激酶选择性,同时减少了脱靶效应,从而减少了副作用。阿氟替尼具有高选择性和强肿瘤缩小特性,对野生型EGFR的抑制作用很小。自2021年3月起,它已在中国被批准用于治疗局部晚期或转移性EGFR敏感突变的NSCLC。莫西替尼(TAK-788)是一种新型口服靶向EGFR/HER2药物,属于第四代EGFR抑制剂。结构上与奥希替尼相似,对EGFR外显子20插入和其他非敏感突变具有增强的抑制作用,对HER2外显子20插入突变的肺癌具有一定的抑制作用。2021年9月,FDA批准莫丝替尼治疗转移性NSCLC、EGFR突变晚期NSCLC、局部晚期NSCLC和晚期NSCLC。该药物还于2023年1月在中国获得上市许可。奥穆替尼(HM61713.BI-1482694)是一种口服有效的小分子,具有潜在的抗肿瘤活性,是韩美制药株式会社开发的用于治疗NSCLC和肺腺癌的突变选择性第三代EGFR抑制剂。它与激酶结构域附近的半胱氨酸残基结合,从而诱导表达EGFR的肿瘤细胞中的细胞死亡。2016年5月获批在韩国上市,用于治疗EGFR阳性的局部晚期或转移性NSCLC患者T790M突变。奈拉替尼是一种口服、有效且不可逆的第三代EGFR TKI,通过阻断泛HER家族(HER1、HER2和HER4)和下游信号通路转导来抑制肿瘤生长和转移。这种药物最初由惠氏(现为辉瑞)和Puma生物技术公司开发。它不仅竞争性地占据EGFR上的ATP结合位点,而且还与靠近口袋开口的独特氨基酸残基Cys805 (EGFR cys797的同源半胱氨酸残基)结合,发生烷基化或共价键,从而实现对HER2的不可逆抑制。奈拉替尼于2017年7月获得FDA批准用于治疗乳腺癌,使其成为世界上唯一获批用于曲妥珠单抗(赫赛汀)强化辅助治疗HER2阳性乳腺癌以降低复发风险的产品。吡咯替尼(SR-1258)是由江苏恒瑞医药有限公司研发的,是一种有效、选择性和不可逆的HER2/EGFR双靶点酪氨酸激酶抑制剂,IC50值分别为38 nM和13 nM。作为第三代EGFR TKI, 吡咯替尼共价结合EGFR、HER2和HER4细胞内激酶区域的ATP结合位点,阻止同型二聚体的形成,从而不可逆地抑制自磷酸化,阻断下游信号通路的激活,抑制肿瘤细胞生长。于2018年8月获得国家药品监督管理局(NMPA)有条件上市批准。3. 共价p53调节剂P53是一种至关重要的蛋白质,它调节细胞周期并发挥肿瘤抑制作用。研究表明,大约一半的人类癌症,包括浆液性卵巢癌、肺鳞状细胞癌、肺小细胞癌、三阴性乳腺癌和鳞状食管癌,p53基因发生改变,导致p53功能丧失或p53表达减少。作为一种与PPIs密切相关的肿瘤抑制因子,p53在调控基因表达、促进肿瘤细胞周期阻滞、细胞凋亡和DNA修复等方面发挥着关键作用。它可以激活附近或远处的基因以响应增强子,同时也间接抑制许多基因的转录。P53可分为突变型和野生型,突变型P53促进肿瘤发生,野生型P53具有广谱肿瘤抑制作用。TP53突变通常会降低p53蛋白的表达或产生无活性变体,从而损害其抑癌特性。因此,需要治疗策略来恢复p53的功能。然而,大多数小分子通过抑制过表达蛋白的活性来靶向过表达蛋白,使p53成为“不可成药”的靶标。直接靶向p53的共价调节剂2022年,Kevan M. Shokat团队开发了p53-Y220C突变体的小分子共价抑制剂KG13。该抑制剂是专门设计用于结合p53 Y220C突变体,使p53蛋白的热稳定性恢复到野生型p53蛋白的水平,激活下游基因的表达。研究人员设计了13种小分子药物,针对p53 Y220C在空间中形成的口袋结构。经过一系列的结构修饰和筛选,KG13被选为最佳的小分子化合物,共价标记率和热稳定性回收率最高。此外,用KG13处理的细胞显示p53 Y220C依赖性p53靶基因激活,抑制细胞生长,增加caspase活性。共价p53-MDM2 PPI抑制剂小鼠MDM2和MDMX都是p53的负调节因子,通过直接结合其N端并介导其在正常细胞中的降解,将p53维持在低表达水平。p53降解的主要机制涉及E3泛素连接酶MDM2的泛素化,从而导致p53的蛋白酶体降解。由于MDM2介导的泛素化和降解依赖于其与p53的直接相互作用,研究人员一直在寻找能够抑制这种相互作用的小分子,以稳定p53并恢复其活性。虽然大多数p53-MDM2抑制剂是非共价的,但一些针对p53-MDM2的小分子抑制剂已被发现是共价的,从而促使了共价p53-MDM2抑制剂的发展。2017年,Ishiba等人通过D-proteins进行镜像筛选,发现NPD6878(阿波啡)是一种高效的MDM2-p53抑制剂候选药物。。在等效剂量下,R-(-)-阿波啡抑制了天然的L-MDM2-L-p53相互作用(IC50 = 0.215 μM)和镜像的D-MDM2-D-p53相互作用(IC50 = 0.195 μM)。此外,对映体S-(-)-阿波啡对L-MDM2-L-p53相互作用也表现出同等的抑制活性(IC50 = 0.175 μM)。2021年,Hamachi等人基于NASA反应基团开发了一种小分子共价抑制剂化合物,可以阻止HDM2与p53的相互作用。他们发现HDM2上的修饰位点是N端α -胺和Tyr67。使用Nutlin-3作为共价抑制剂的支架,NASA弹头可以靶向的Lys51与Nutlin-3a的2-氧嘧啶部分的距离约为11 Å。然后对其进行结构修饰,生成一系列共价化合物,并对其在体外修饰HDM2的能力进行了测试。4.其他共价抑制剂共价Mcl-1抑制剂Mcl-1是Bcl-2蛋白家族中一个重要的抗凋亡成员,在各种人类癌症的发展中起着重要作用。靶向Mcl-1的BH3结合槽是抑制其功能的一种很有前景的方法,已成为抗肿瘤药物开发的热点。在这方面,Lee等人设计了一种基于与结合位点相对的BH3区域附近可变结构位点表的药物设计策略。他们利用共价抑制剂MAIM1与thalproquinone型Cys286结合,有效抑制Mcl-1活性。该化合物与Mcl-1紧密结合,为抗凋亡肿瘤治疗提供了潜在的新途径和药物前体化合物。共价PKM2抑制剂在最近一项关于靶向共价抑制剂的研究中,上海有机科学研究所报道了一种基于三价砷共价弹头的新型PKM2抑制剂化合物。三价砷官能团具有作为共价弹头的潜力,因为它能够与蛋白质中的半胱氨酸残基反应,影响其活性并发挥药物作用。利用有机砷共价探针、化学蛋白质组学和药物化学方法,开发了一种高活性和特异性的共价PKM2抑制剂,该化合物在体内有效抑制卵巢癌的生长。02变构调节剂合理药物设计的最初重点是治疗性蛋白靶点的正构位。然而,由于它们与底物的高亲和力,缺乏结构信息或活性位点的高度保守性,许多这些靶点被发现是不可成药或难以靶向其正构位。为了克服这些挑战,变构调节已被提出作为一种在自然界中常用的策略,通过“远距离”调节生物分子的亲和力来控制细胞过程。变构调节剂可以通过将靶蛋白稳定在非活性或活性状态,以高度可预测的方式改变蛋白质/底物亲和力,从而实现理想的可控性。变构调节剂比正构抑制剂有几个优点。首先,变构配体不必与高亲和力底物竞争,这使得开发变构调节剂变得更简单。其次,变构位点具有多样性,对同源蛋白具有更好的选择性,副作用更小,具有更大的临床应用价值。第三,变构调制器具有理想的“效应上限”。一旦变构位点被占据,没有观察到额外的效应,表明药物在过量条件下是安全的,此外,不可成药的蛋白质可以同时被正构抑制剂和变构调节剂靶向,以实现协同效应并克服耐药性。变构调节剂不仅可以像正构抑制剂一样抑制靶标,还可以在需要时稳定或竞争性地占据它们以改善病理状态。根据对受体的作用,可将变构调节剂分为三类:正变构调节剂(positive allosteric modulators, PAMs),它改善了正构效应剂的作用,但没有内在活性;负变构调制器(NAMs),抑制正构效应器的功能;沉默变构调节剂(SAMs),也称为中性变构调节剂,通过阻断PAM和NAMs的变构位点来抑制变构活性。图2 针对不可成药蛋白质的变构抑制剂1.RAS变构抑制剂KRAS-GTP或KRAS-GDP的相互作用与KRAS的激活状态密切相关,进而影响其信号传输。一般来说,参与KRAS- gdp结合的非活性KRAS构象是首选。。结构生物学分析鉴定出KRAS蛋白表面的两个开关,开关I和开关II。开关II区是特别重要的,因为它的高构象变异性,这为变构调节提供了切入点。与开关II区突变氨基酸相互作用的变构抑制剂可以诱导构象变化,导致更不活跃的KRAS构象。因此,开发特异性靶向KRAS并抑制其异常功能的变构抑制剂为靶向KRAS突变体提供了一种很有前景的方法。KRASG12C变构抑制剂KRASG12C突变蛋白含有突变型半胱氨酸Cys12,它为抑制剂提供了一个潜在的共价位点。当带有共价弹头的抑制剂与突变体Cys12共价结合时,它在开关II区诱导一个新的变构口袋S-II P。然后,小分子抑制剂与相应的氨基酸相互作用,导致KRAS蛋白的构象变化。这些作用降低了GTP和KRAS之间的亲和力,阻止GTP通过GEF催化取代GDP,最终将KRAS突变体锁定在非活性状态。基于这一作用位点,已经开发出几种靶向KRAS的小分子抑制剂,这些药物包括Sotorasib (AMG-510)、Adagrasib (MRTX-849)、ARS-853、ARS-1620、LY-3537982、GDC-6036 (RG-6300)、D-1553、ARS-3248 (JNJ-74699157)、JDQ-443和SML系列化合物,以及LY-3537982、ARS-853、ARS-1620和6H05系列化合物。KRASG12D变构抑制剂KRASG12D是一种更普遍的KRAS突变类型,在各种癌症中都有发现,包括胰腺癌、结直肠癌和肺腺癌。因此,它是开发选择性KRAS突变抑制剂的潜在靶点。然而,与KRASG12C不同,KRASG12D在开关II结合袋附近缺乏活性残基,这阻止了蛋白质进行共价修饰。Mirati
Therapeutics已经确定并描述了一种选择性、非共价、高亲和力的KRASG12D抑制剂,称为MRTX-1133。该抑制剂与KRASG12D的失活形式结合,IC50 < 2 nM。MRTX-1133还抑制RAF-RAS结合域肽与KRASG12D活性形式的结合,IC50为9 nM,并诱导KRAS蛋白开关I和开关II区域的构象变化。MRTX-1133通过在KRAS蛋白开关II区与天冬氨酸Asp12和谷氨酸Glu62相互作用,发挥变构作用,导致KRAS蛋白构象改变,抑制KRASG12D突变细胞和肿瘤环境中KRAS信号通路,从而达到抗肿瘤作用。SHP2变构抑制剂SHP2是一种由PTPN11基因编码的非受体蛋白酪氨酸磷酸酶(PTP)。SHP2参与多种信号通路的调控,包括与癌细胞相关的RAS-MAPK、PI3K-AKT和JAK-STAT通路。此外,SHP2还与PD-1/PD-L1的某些功能有关,从而起到调节免疫系统的作用。此外,SHP2过表达或激活可介导多种癌症的耐药,包括白血病、非小细胞癌和乳腺癌。因此,SHP2被认为是癌症治疗的潜在靶点。在结构上,SHP2在N端包含两个SH2结构域(N-SH2和C-SH2),一个催化PTP结构域,在C端包含两个可磷酸化的酪氨酸残基(Tyr542和Tyr580)。通常,N-SH2结构域和PTP结构域之间的相互作用导致SHP2蛋白的自动抑制封闭构象。在生长因子或细胞因子的刺激下,SHP2蛋白被激活,由于占据SH2结构域从而阻断N-SH2-PTP相互作用,从而暴露出催化PTP结构域。因此,SHP2的催化位点对其底物可用,随后的信号转导可以被激活。最初,调控SHP2的尝试主要集中在识别PTP结构域特有的传统竞争性抑制剂,也称为正位抑制剂。然而,这些正构抑制剂对高度同源的蛋白(SHP1, SHP2和PTP1B)选择性低,毒性大且细胞通透性低,从而导致生物利用度差。因此,SHP2长期以来一直被认为是一种“不可成药”的靶蛋白。变构调节剂的工作原理是直接稳定SHP2的自抑制构象,从而阻止催化PTP结构域与SHP2底物之间的相互作用。在过去的几十年里,通过高通量筛选已经发现了许多这种类型的SHP2抑制剂,包括TNO-155,JNB-3068,JAB-3312,RLY-1971等。这些抑制剂中有许多已经进入临床试验阶段,但目前都还没有被批准用于临床。GPCR变构调节剂G蛋白偶联受体(GPCR在人体生理中起着重要作用,是市场上许多药物的主要靶点。虽然最初的努力集中在开发靶向活性位点内源性配体(正位位点)的正位抑制剂上,但这些抑制剂的选择性受到GPCR中正位结合位点高度保守性的阻碍。因此,通过设计基于不同变构口袋的不同亚型联合,或与正构调节剂联合,变构调节剂为控制GPCR活性开辟了新的可能性。基于在GPCR中发现的变构位点,以及基于配体或结构的变构药物的发现和设计,已经批准上市了多种相关药物,包括Avacopan, Cinacalcet, Ticagrelor和 Maraviroc。还有数十种分子正在I-III期临床试验中进行测试。03PPI抑制蛋白-蛋白相互作用(PPIs)异常可导致各种疾病,具有平坦活性位点的不可成药蛋白质通常通过PPI网络起作用。迄今为止,已经开发出多种针对PPI的调节剂,其中许多已成功进入临床。就结构类型而言,现有的PPI抑制剂可分为几类,包括小分子、抗体、肽或重组蛋白。“Hot spots”的发现使得有效地靶向PPIs成为可能。在蛋白质之间形成结合位点的和hot spot残基称为PPI界面。根据抑制剂是否与PPI界面结合,PPI调节剂可分为正构抑制剂和变构调节剂。正构抑制剂通过识别hot spot来靶向PPI界面,而变构调节剂通过调节靶蛋白的非相互作用区域来诱导靶蛋白的构象变化。同样,根据结合方式,PPI调节剂可分为非共价调节剂和共价调节剂。图3 靶向不可成药蛋白的PPI抑制剂和蛋白质- DNA相互作用抑制剂1.RAS - PPI抑制剂RAS-RAF 抑制剂基于合成蛋白(FN3)骨架的单体蛋白在功能上与抗体相似,可以与相应的靶标结合使用。与传统抗体相比,这些蛋白质具有更小的分子量和更清晰的结构。此外,它们在氧化/还原条件下更稳定,使其成为新型的理想的抑制剂。研究人员利用单体蛋白文库发现了一种特异性结合HRAS而非NRAS的单体蛋白NS-1。NS-1和HRAS的结合位点位于RAS/RAF界面对面的RAS蛋白α4、α5和β6区。NS-1的结合可以变构诱导SW-I和SW-II结构域的结构变化,从而破坏RAS与包括RAF在内的效应蛋白之间的相互作用。细胞实验表明,NS-1能有效抑制细胞中RAS蛋白下游MAPK信号通路的激活。在膀胱癌细胞中的实验表明,NS-1能够抑制内源性突变的RAS信号和相应的细胞增殖。这些结果表明,通过变构效应破坏RAS与RAF之间的蛋白相互作用可以抑制RAS蛋白介导的癌变,为开发针对RAS蛋白的抗癌疗法提供了新的思路。RAS-SOS抑制剂SOS是一种鸟嘌呤核苷酸交换因子(GEF),对细胞中KRAS蛋白的激活至关重要,与KRAS及其下游信号通路的表达密切相关。RAS- SOS复合物改变RAS蛋白的构象,阻断镁辅因子与GDP磷酸基的亲水性相互作用,从而降低RAS对GDP的亲和力,促进其释放。Boehringer
Ingelheim(BI)开发了BI-1701963,这是第一个泛KRAS抑制剂,它对KRAS等位基因(G12D, G12V, G12C和G13D)具有广泛的活性,同时保留了KRAS- SOS2的相互作用。BI-1701963在多种疾病的治疗中具有潜在的应用价值,包括腺癌、晚期实体瘤、胆道癌、结直肠癌、肺癌、转移性结直肠癌和转移性胰腺癌。RAS-PDEδ 抑制剂KRAS蛋白位于细胞膜内侧,通过法尼基修饰基团附着在细胞膜上。在法尼基转移酶的作用下,通过翻译后蛋白修饰将法尼基添加到KRAS蛋白上。KRAS只有位于细胞膜上时才能被上游信号蛋白完全激活因此,抑制法尼基转移酶的活性可以阻止KRAS的法尼基化,减少其在细胞膜上的定位,最终抑制KRAS的活性。PDEδ是一种结合KRAS蛋白并调节其向膜室方向的蛋白。PDEδ具有一个大的疏水口袋,可以结合法尼基蛋白,特别是KRAS蛋白的脂质部分,从而阻止法尼基修饰的KRAS蛋白与细胞膜结合,抑制肿瘤细胞的增殖和生长。通过分子对接和高通量虚拟筛选,开发了多种KRAS-PDEδ抑制剂,包括三唑抑制剂化合物、四氢二苯并呋喃抑制剂NHTD和香豆素抑制剂Deltaflexin。其中,三唑类PDEδ抑制剂有效阻断了PDEδ与RAS蛋白之间的相互作用。另一方面,NHTD通过PDEδ靶向异戊二烯结合袋,将KRAS的定位重新分配到子宫内膜。Deltaflexin选择性地破坏KRAS,同时保留HRAS的膜结构。此外,Deltaflexin选择性抑制致癌KRAS驱动的细胞增殖和肿瘤形成。2.Bcl-2 PPI抑制剂Bcl-2是一种抗凋亡蛋白,在多种血液系统恶性肿瘤中高表达,使其成为一个有吸引力的药物靶点。促凋亡蛋白如Bax和Bad对细胞凋亡过程至关重要,但当它们与抗凋亡蛋白如Bcl-2结合时,其功能受到抑制。因此,阻断促凋亡和抗凋亡蛋白之间的相互作用可以阻止肿瘤细胞逃避凋亡。Bcl-2与Bak/Bax的PPI界面面积约为750-1500 Å2,较为平坦,缺少凹槽。目前已开发出多种Bcl-Bax抑制剂,包括ABT-199
(venetoclax)、ABT-737、ABT-263
(navitoclax)等。其中ABT-199是首个也是目前唯一获批的小分子Bcl-2抑制剂。研究人员在ABT-263的骨架结构中引入吲哚基和偶氮吲哚基,合成ABT-199。ABT-199已被FDA批准用于治疗成人慢性淋巴细胞淋巴瘤(CLL)或小淋巴母细胞淋巴瘤(SLL)。临床试验表明,ABT-199联合抗CD20单克隆抗体利妥昔单抗比ABT-199单药治疗更有效。3. P53-MDM2 PPI 抑制剂MDM2是p53最重要的负调节因子,直接与p53结合并形成复合物,调节p53蛋白的稳定性和活性。MDM2结合p53转录激活域的N端,抑制p53的反式激活和细胞生长,调节细胞周期,诱导细胞凋亡。因此,破坏MDM2-p53相互作用,提供了一种通过恢复p53受损功能来治疗癌症的潜在方法。目前开发出的MDM2-p53抑制剂包括RG-7112,JNJ-26854165,MK-8242
(SCH900242),SAR-405838,AMG-232,NVP-CGM097,DS-3032,NVP-HDM201等。4. Myc PPI 抑制剂Myc癌基因,包括它的三个亚型C-Myc、N-Myc和L-Myc,被认为是一个转录“放大器”,可以在不同类型的癌症中触发各种致癌转录程序。Myc的过表达或过度激活是癌症中最常见的现象之一,使其成为肿瘤发生的关键因素。Myc的大而平坦的相互作用表面形成蛋白质复合物,使其成为药物开发的困难靶点。因此,Myc被认为是一个无法成药的靶标。到目前为止,还没有针对Myc的药物被批准上市。近年来,作用于Myc-PPI的抑制剂的开发取得了重大进展,包括三种主要类型:Myc-Max PPI抑制剂、Myc-WDR5 PPI抑制剂和Myc-TRRAP PPI抑制剂。Myc-Max PPI 抑制剂C-Myc通过bHLH-ZIP结构域与Max形成复合体,激活基因转录。Myc-Max复合物具有两种调控机制:(1)Myc-Max结合启动子区域的特定识别位点(E-Box元件)激活基因转录;(2) Myc- max异源二聚体通过锌指蛋白mz -1间接募集到DNA中,从而抑制Myc调控基因。考虑到这两种调节机制都依赖于Myc-Max相互作用,调节这种相互作用是抑制Myc功能的有效方法。Omomyc是研究最多的c-Myc/Max抑制剂之一,它与c-Myc竞争与Max的结合,形成Omomyc/Max复合物,阻止参与增殖和代谢的Myc靶基因的转录激活。另一方面,Max-Max同型二聚体的形成为阻断Myc/Max相互作用和阻止Myc/Max异源二聚体与DNA结合提供了另一种机制。稳定Max-Max同型二聚体可以防止Myc-Max靶基因的反激活。一些活性小分子已被确定为有前途的Max-Max同型二聚体稳定剂,包括NSC13728和KI-MS2-008。Myc-WDR5 PPI 抑制剂WDR5是一种高度保守的WD-40重复蛋白,在造血过程中发挥关键作用,调节H3K4甲基化和HOX基因表达。Myc-WDR5相互作用是通过调控基因表达来控制细胞增殖、分化和凋亡的关键。Myc通常与WDR5的WBM位点结合,WDR5在Myc蛋白中具有明显的保守结构特征,因此非常适合开发Myc-WDR5的小分子抑制剂。2019年,Macdonald等人通过高通量筛选发现了一类具有磺胺结构的Myc-WDR5抑制剂。虽然化合物(170)的活性最高(IC50 =
29 nM),但不适合进行体内研究。为了解决这个问题,Chacon Simon等人通过核磁共振片段筛选鉴定了其他几个化合物片段,得到化合物(171)(IC50 =
100 nM)该化合物目前被用于研究Myc-WDR5复合物的生物学效应。Myc-TRRAP PPI抑制剂TRRAP是一种高度保守的434 kDa蛋白,属于PI3K家族。它是Myc的关键辅助因子和组蛋白乙酰化(HAT)复合体的成员。Myc和TRRAP之间的相互作用发生在Myc蛋白的一个称为MB-II的精确区域,该区域也是Myc转激活结构域(TAD)的核心。MB-II在与TRRAP相互作用时成为一个明确定义的结构,这为开发能够阻断Myc-TRRAP相互作用并特异性靶向Myc驱动的癌症的抑制剂提供了机会。2021年,Alexander等人发现姜黄素干扰Myc的基本功能,并诱导这种致癌TF与其共激活因子TRRAP之间的交联。5. TF–DNA 相互作用抑制剂除了PPI抑制剂外,阻断TF与DNA的相互作用是另一种选择。蛋白质- DNA的作用界面直接暴露在溶剂中。因此,具有高正电荷的氨基酸残基,如赖氨酸和精氨酸,可以直接与DNA骨架匹配。在过去的二十年里,在开发能够特异性地与DNA结合以调节TF活性的小分子方面取得了重大进展。例如,Hiroshi Sugiyama的团队最近开发了一种基于吡咯-咪唑聚酰胺的新型人工TF模拟化学物质,可以靶向特定的DNA序列并调节基因表达;Carabet等人利用MOE的Site Finder模块,根据与DNA序列结合的c-Myc-Max异二聚体的晶体结构,在Myc-Max-DNA界面上识别潜在的结合位点。最终得到两种化合物,VPC-70067(191)和VPC-70063(192);2022年,Artem Cherkasov的研究小组在之前的研究中发现了Myc bHLHZ结构域的口袋,从而发现了化合物VPC-70063作为c-Myc抑制剂。通过一系列分子对接虚拟筛选和优化,他们随后获得了化合物VPC-70619(193),这是一种有效的口服活性n-Myc抑制剂,与DNA结合区结合,阻止N-Myc/Max与DNA结合;2022年,徐克林课题组根据c-Myc/Max配合物的晶体结构,在化合物文库中进行分子对接筛选,鉴定出活性最高的化合物D347-2761(194)。D347-2761与Myc蛋白的DNA结合区结合,在5 ~ 10 μM范围内对骨髓瘤细胞表现出增殖抑制活性。04靶蛋白调控蛋白质调控可分为两大类:蛋白质降解和蛋白质稳定。近年来,通过靶向蛋白水解嵌合体(proteolysis-targeting chimeras, PROTAC)直接降解蛋白质治疗疾病的显著进展凸显了蛋白质调节药物的重要性。图4 靶向蛋白调控策略针对不可成蛋白1. 基于蛋白酶体的靶向蛋白降解(TPD)PROTAC作用机制的核心是通过泛素-蛋白酶体系统(UPS)降解靶蛋白。作为一种异双功能分子,PROTAC同时靶向目标蛋白和E3泛素连接酶,形成三元配合物。在这个复合体中,目标蛋白被泛素标记,促进其被蛋白酶体识别和随后的降解。与传统的小分子抑制剂相比,PROTAC采用“事件驱动”的降解过程,而不是“占用驱动”的结合机制,使PROTAC高效并能够快速降解细胞内靶蛋白,包括耐药突变体。PROTAC技术具有降解近80%人类细胞内蛋白的潜力,在各个领域都有广泛的应用,最显著的是在癌症治疗中。图5 针对不可成药蛋白的靶向蛋白调节剂的选定化学结构2. 靶向蛋白质降解的分子胶与PROTACs不同,分子胶具有E3泛素连接酶和靶蛋白的双配体结构,能够泛素化和降解以前不可成药的靶标和PPIs。分子胶的特点是分子量更小,化学机制更简单,空间干扰更少,药物形成更优越。然而,它们不能像PROTACs那样通过大规模的组件筛选来设计。分子胶方法有效地克服了传统抑制剂的局限性,使以前“不可成药”的靶标变得“可成药”。3.其他基于蛋白酶体的TPD药物研究技术p-PROTAC多肽以其独特的物理化学性质,弥合了小分子和大分子之间的差距,使小分子药物能够进入具有挑战性的药物结合位点,并有助于减轻与小分子药物相关的耐药性。在2020年,Chen的团队报告了一种基于肽的PROTAC—xStAx-VHL,,来降解β-连环蛋白。该PROTAC直接识别该蛋白并通过泛素-蛋白酶体途径促进其降解。PROTABPROTABs蛋白降解技术平台采用双特异性抗体方法。它包括一个靶向跨膜E3连接酶的泛素连接酶的结合域(anti-tag),如RNF43或ZNRF3,以及一个靶向蛋白的结合域(anti-POI)。在一项研究中,PROTAB有效地降解了直肠癌患者的胰岛素样生长因子1受体(IGF1R)。研究人员还通过降解Her2和PD-L1靶点证明了该平台的潜力。CHAMP伴侣介导的蛋白质降解(CHAMP)是利用分子伴侣的另一种基于蛋白酶体的蛋白质降解技术。CHAMP的结构类似于PROTAC,一端是靶向蛋白配体,另一端是分子伴侣,由合适的连接体连接。与PROTAC一样,CHAMP促进靶蛋白的泛素化及其随后被蛋白酶体降解。Lysosome-based
TPD溶酶体和蛋白酶体系统作为细胞内两种主要的蛋白质降解途径,具有不同的功能。蛋白酶体系统主要降解短寿命、可溶性和错误折叠的单体蛋白质,而溶酶体系统主要降解长寿命蛋白质、蛋白质聚集体和受损的细胞器。这两个系统都可以通过泛素化实现底物降解,也可以协同降解相同的细胞内底物。利用溶酶体途径进行蛋白质降解允许靶向更广泛的底物,包括传统上不可靶向的蛋白质。该领域目前主要包括两类技术:自噬-溶酶体途径(如AUTACs、ATTEC和AUTOTACs)和内吞-溶酶体途径(如LYTACs和MoDE-As)。4.靶向蛋白稳定(TPS)Nomura的团队受到PROTAC的启发,开发了针对TPS的去泛素酶靶向嵌合体(DUBTAC)。他们将EN523(一种选择性结合OTUB1酶的分子)与lumacaftor结合,产生了NJH-2-057。与单独使用lumacaftor相比,DUBTAC有效抑制ΔF508-CFTR降解,提高蛋白水平,并在囊性纤维化供体细胞中表现出更好的稳定性。在此基础上,Wei的团队开发了一种TF-DUBTAC平台,能够选择性地稳定癌蛋白,包括FOXO3A、p53和IRF3。他们选择Foxo特异性DNA基序作为结合配体,并在DNA基序的5 '端加入叠氮基团。利用Click反应,他们将EN523-BCN和N3-FOXO-ODN连接起来,生成TF-DUBTAC。05基于核酸的方法随着人们越来越认识到RNA在传递细胞信息和调节基因方面所起的重要作用,靶向RNA已成为一个令人兴奋的机会,可以治疗性地调节与以前“不可成药”蛋白质靶点相关的细胞过程。在Watson-Crick互补结合规则的驱动下,可以在遗传/转录水平上调节“不可成药”的蛋白质,这扩大了可成药靶标的范围。基于RNA的治疗方法也被称为寡核苷酸治疗方法,分为反义寡核苷酸(ASOs), RNA干扰(RNAi),包括小干扰RNA (siRNA)和microRNAs (miRNAs),基于CRISPR的基因组编辑和G4稳定。前两种类型在临床发展和治疗应用中最具代表性。在不可成药蛋白质中,siRNA分子在药物发现中占有突出地位。图6 基于核酸的方法针对不可成药蛋白06免疫疗法免疫疗法近年来获得了相当大的关注,因为它在治疗由不可成药蛋白质引起的各种疾病方面具有潜力。免疫检查点抑制剂、疫苗、特异性抗体和过继细胞疗法,已被用于改善与p53和RAS相关的疾病。这些进步为疾病治疗的未来提供了巨大的希望,并强调了在该领域继续研究的重要性。图7 免疫治疗方法针对不可成药蛋白07其他方法1. 针对上游/下游效应物或辅因子由于一些不可成药蛋白靶点涉及复杂的生物网络和信号通路,通过靶向其上游或下游蛋白来调节其相关信号通路已成为间接靶向不可药物蛋白的另一种方法。以RAS为例,目前已确定的RAS效应家族超过11个。RAS上游受体酪氨酸激酶如EGFR家族的失活可降低RAS的活化,抑制EGFR成为治疗RAS突变肿瘤的有效途径。另一方面,MAPK和PI3K是分别可被RAS激活的效应通路,完全抑制MAPK通路有助于治疗RAS突变型肿瘤。因此,RAF抑制剂、MEK抑制剂和ERK抑制剂可与其他抑制剂联合使用,间接靶向突变型RAS。此外,靶向TF是一种很有希望的调节转录失调的方法,因为它们的作用与一般转录辅助因子(如转录激酶、表观遗传蛋白和共激活因子)的化学扰动高度相关。2. 诱导合成致死合成致死指的是两种基因同时突变导致细胞死亡,但任何一种基因单独突变都与生存能力相容的一种病理机制。因此,鉴定突变基因驱动的不可成药蛋白合成致死基因具有重要意义。例如,分别编码CRAF和SHOC2的RAF1和SHOC2已被证明在KRAS突变细胞系中起重要作用,这表明抑制CRAF和SHOC2可用于KRAS突变型癌症。3. 靶向翻译后修饰(PTMs)多种翻译后修饰(PTMs),如泛素化、羟基化、甲基化、乙酰化和磷酸化,在协调不可成药TF的活性中起着关键作用,包括蛋白质稳定性、亚细胞定位、PPIs和序列特异性DNA结合。例如,可以使用PHD抑制剂来抑制HIF-羟基化,例如在中国和欧洲被批准用于治疗CKD肾性贫血的Roxadustat。4. 设计偶联物通过将各种类型的分子(包括抗体、药物和小分子)结合成一个单一的化学实体,已经开发出多种偶联物以结合所需的特性,例如有效的传递功能和细胞毒性活性。这为合理设计靶向不可药物蛋白的潜在化合物提供了一种有吸引力的方法。各种类型的偶联物,包括抗体-药物偶联物(ADC),抗体- siRNA偶联物(ARC),小分子-药物偶联物(SMDC)和小分子辅助受体靶向(SMART),已被设计为不可成药蛋白的抑制剂。特别是ADC,已经成为一种主流的偶联物,已经成功地用于癌症治疗。小结随着尖端药物发现技术的不断进步,人们开发了多种直接或间接靶向不可成药蛋白的方法,包括共价抑制剂的开发、变构位点的鉴定、靶向PPIs、影响蛋白稳定性、调节RNA、调节免疫等。希望随着未来对发病机制的深入研究和更优治疗策略的制定,药物靶点的范围可以继续扩大,开发出更多有前景的药物治疗各种疾病。参考文献:1.Xie, X., Yu, T., Li, X., Zhang, N., Foster, L. J., Peng, C., Huang, W.,
& He, G. (2023). Recent advances in targeting the "undruggable"
proteins: from drug discovery to clinical trials. Signal transduction and
targeted therapy, 8(1), 335. https://doi-org.libproxy1.nus.edu.sg/10.1038/s41392-023-01589-z2. Lu, Y., Yang, Y., Zhu, G., Zeng, H., Fan, Y., Guo, F., Xu, D., Wang, B.,
Chen, D., & Ge, G. (2023). Emerging Pharmacotherapeutic Strategies to
Overcome Undruggable Proteins in Cancer. International journal of biological
sciences, 19(11), 3360–3382. https://doi-org.libproxy1.nus.edu.sg/10.7150/ijbs.83026作者:一颗草莓推文用于传递知识,如因版权等有疑问,请于本文刊发30日内联系医药速览。原创内容未经授权,禁止转载至其他平台。有问题可发邮件至yong_wang@pku.edu.cn获取更多信息。©2021 医药速览 保留所有权利关注我,更多资讯早知道↓↓↓