更新于:2024-09-19
Zhejiang Dongfang Shaoxing Wine Co. Ltd.
更新于:2024-09-19
概览
关联
100 项与 浙江东方绍兴酒有限公司 相关的临床结果
登录后查看更多信息
0 项与 浙江东方绍兴酒有限公司 相关的专利(医药)
登录后查看更多信息
20
项与 浙江东方绍兴酒有限公司 相关的新闻(医药)2024-09-02
8月31日,2024年抗肿瘤药物创新研发大会暨《2023年度中国抗肿瘤新药临床研究评述》发布会,在上海隆重举行。本次大会由中国医药创新促进会抗肿瘤药物临床研究专业委员会(以下称“抗肿瘤专委会”)、艾美达医药咨询、北京市希思科临床肿瘤研究基金会以及东方临床肿瘤研究中心联合主办。大会重点围绕“抗肿瘤领域源头创新”与“新形势下抗肿瘤新药研发的挑战与出路”两大主题展开报告分享与讨论,吸引了众多业内临床专家、学者及企业代表的参与。
年度报告重磅发布
中国医药创新促进会执行会长宋瑞霖与抗肿瘤专委会主任委员、中国药科大学附属上海高博肿瘤医院院长李进分别致开幕词。
宋会长指出,《2023年度中国抗肿瘤新药临床研究评述》(下称“《评述》”)的发布将为未来抗肿瘤药物研发提供科学依据,为产业发展指明了方向。宋会长呼吁在循证医学转向精准医学的当下,新药研发更需医、研、企的紧密合作。
李进院长表示此次大会是响应国家“2030健康中国”战略目标的具体行动。他特别提及了《评述》的编写过程,并表示该书是对抗肿瘤领域新药研发形势的重要总结,希望《评述》的发布能够为中国新药创制的发展贡献积极力量。
抗肿瘤专委会秘书长薛俊丽主持第一阶段会议,并详细介绍了《评述》的编撰过程。她回顾了过去几年中,抗肿瘤专委会在推动新药临床研究发展方面所做出的工作。同时薛秘书长表示《评述》不仅涵盖了当前热门靶点的深入分析,还对未来的研发方向提供了战略性建议,希望能够帮助研究者和企业在新药开发中更好地把握机会、优化资源配置。
在宋瑞霖会长、冯岚秘书长、李进院长、马军教授、周彩存主任、韩宝惠院长、赵洪云主任、刘爱军主任以及薛俊丽秘书长的共同见证下,《2023年度中国抗肿瘤新药临床研究评述》正式发布。
《2023年度中国抗肿瘤新药临床研究评述》.pdf
多方发力、共建源头创新
第二阶段会议由中国药促会抗肿瘤临床专委会常务委员,上海市胸科医院副院长、主任韩宝惠教授主持。
李进院长分享了“中国抗肿瘤药物研发的未来展望”主题报告,深入分析了当前中国抗肿瘤药物研发的复杂局面,并指出企业家和政府需共同努力提升中国生物制药行业的全球竞争力。他提出了利用商保支持、跨境数据传输等实际解决方案,推动中国新药创制的国际化进程。
在主题讨论环节,马军教授作为主持人与周彩存主任、韩宝惠院长、蔺强院长、赵洪云主任以及圣和药业产品与创新体系部门负责人李佳妮博士围绕“中国抗肿瘤领域源头创新发展之路”展开讨论。
嘉宾们一致认为,当前中国新药研发源头创新的缺失主要在于企业创新研发与临床结合的紧密度不足。建议进一步发挥临床在药物研发链条中的重要作用,尤其是在药物早期研究与耐药机制研究中,同时建议企业增加在转化研究和精准医疗方面的投入。嘉宾们一致表示中国医药创新的高质量发展必须建立在协作、转化与科学决策的基础上,才能在全球竞争中脱颖而出。
抗肿瘤新药研发:新技术、新形势、新挑战
第三阶段会议由中国医药创新促进会秘书长冯岚主持。
抗肿瘤专委会副主任委员、同济大学附属东方医院肿瘤科主任周彩存以“肺癌治疗与药物研发新趋势”为主题进行报告分享。周主任强调新药临床研发的方案选择应与精准医学相结合、与诊疗实际相结合,对照药应选择实际临床诊疗中的最佳治疗方案。周主任提出中国肺癌患者的不同驱动基因的比例与美国存在较大差别,药物创新研发不能盲目跟随美国,需要从自身出发,研制更适合中国人的中国药。
抗肿瘤专委会副主任委员、哈尔滨血液病肿瘤研究所所长马军进行了题为“血液淋巴系统疾病治疗及研发的回顾和进展”的报告分享。他指出尽管在CAR-T等先进疗法的不断涌现下淋巴瘤的治疗已经取得了显著进展,但仍然面临T细胞靶向治疗复杂性和二次癌症风险等挑战。他特别提到,当前的医学研究不仅要关注新药研发的创新性,还要充分考虑药物的安全性和患者的长期生存质量。
因美纳全球高级副总裁兼大中华区总经理郑磊以“基因测序技术助力全球源头创新之中国战略与思考”为主题作报告分享。她指出,基因测序已成为精准医疗的重要基础,通过大数据和AI技术可以进一步提升药物研发成功率。她倡导推动生物样本库的应用和产业转化,为未来的技术创新和药物开发奠定坚实基础。
IQVIA大中华区商务拓展副总裁郭彤围绕“抗肿瘤临床试验数字化管理进程”做主题报告。他系统地介绍了全球数字化临床试验的三大发展阶段,并指出临床试验数字化的核心在于提高效率、降低成本和提升数据质量。此外,他还探讨了去中心化临床试验在全球和中国的应用案例,展示了数字化工具如何在远程访视、数据采集和患者管理中发挥作用,为未来的临床试验提供了更加灵活和高效的解决方案。
第二场圆桌讨论“新形势下抗肿瘤新药研发面临挑战与出路”由李进院长主持,与蔡清清主任、刘爱军主任、薛俊丽主任及海昶生物医药技术有限公司首席医学官门宇欣围绕临床试验的设计、执行和质量控制等问题分享了宝贵的经验和见解。
嘉宾们从各自研究领域出发,从临床需求程度与药物差异化创新等角度对药品附条件批准上市的可行性进行深入讨论。同时讨论嘉宾一致强调了“沟通”在临床试验中的重要作用:临床试验方案保持科学性需要申办方与研究者加强沟通;降低患者脱落率需要研究者与患者的良好沟通;全球多中心临床试验的高效率推进需要多方加强沟通合作以妥善解决各国的文化和医学差异。
李进院长为大会作总结发言。李院长表示本次大会展现了当前抗肿瘤药物研发正迈向创新与挑战并存的阶段,所有报告与讨论均指向了一个共同的主题——如何在严峻的市场与监管环境中,通过科学、精细化的管理和创新,提升药物研发的成功率。这些探讨将为中国乃至全球的抗肿瘤药物创新提供宝贵的经验与启示,激励着我们在不断前行的道路上,为攻克癌症贡献更多力量。
点击“阅读原文”查看更多报告
核酸药物
2024-08-24
“BLA杯”征文投稿
作者:赵宝龙
来源:蒲公英Ouryao
1
一个人
在人类历史的长河里,疾病如同暗夜幽灵,悄无声息地夺走生命的光辉。脊髓灰质炎,这个名字在20世纪初期,如同一道阴影笼罩全球。它以高度传染性和致命的神经系统攻击性,令无数家庭陷入绝望。1952年,美国57628例患者的记录,数字惊人。而在东方,中国亦未能幸免。1955年,江苏南通市的疫情,如惊雷劈空,震惊全国:1680名儿童患病,28%的死亡率,70%的患者面临瘫痪的残酷现实。
在中国脊灰疫苗问世之前,相关研究尚处于初级阶段。虽然国际上有“灭活疫苗”和“减毒活疫苗”两种技术路径,但受当时复杂国际环境所困,中国无法依赖外国技术。因此,成为了守护国民健康的必由之路。
在资源匮乏、技术封锁的艰难条件下,他选择了成本效益更高的减毒活疫苗技术路线,并在云南昆明建立了庞大的疫苗生产基地。他身先士卒,亲试疫苗,甚至瞒着家人,将这份勇气与牺牲带给自己的孩子,为疫苗的安全性提供了最坚实的证明。
最终,糖丸疫苗的诞生,不仅解决了疫苗储存与运输的难题,更成为了中国公共卫生史上的一座里程碑,阻断了脊髓灰质炎的本土传播,2000年,中国宣告无脊灰。
他,是顾方舟。
2
一家企业
生物制药工艺耗材主要包含一次性工艺袋、色谱填料/层析介质、培养基及除病毒过滤。其中,一次性生物工艺薄膜,以二三十年的历史,见证生物药品生产的演进。它以灵活性、便利性、安全性,在细胞培养、产品混合、储存转移中扮演关键角色。大规模细胞培养技术与一次性生物反应器的发展,更使其应用扩展至抗体、疫苗生产,逐步替代传统不锈钢系统。
根据市场调研机构的数据,2021至2022年间全球市场容量约为82至122亿美元,年复合增速约16%至22%。国内一次性袋子市场在2022年大约为50至60亿元人民币,年复合增速13%至20%。
然而,在这片充满机遇的蓝海中,“蛋糕”虽大,全球市场的主导权却依旧掌握在Cytiva、Sartorius、Thermo Fisher、Merck等国际巨头手中。薄膜材料的自主研发,成为了制约国内行业发展的瓶颈。
2011年,上海一隅,一家默默无闻的小公司悄然成立。两层工厂,四面楚歌,无人问津的窘境中,它面临着人才、技术、资金、市场的多重困境。然而,十年磨一剑,2021年,它的产品已经覆盖了一次性袋子及配套产品、一次性使用设备系统等多个领域,市场份额高达20%-30%的佳绩,向世界证明了自己的实力。
LeKrius®一次性生物工艺薄膜的诞生,打破了国外供应商的垄断,成为国内生物药耗材领域的领军者。
它,是乐纯生物。
3
一个行业
自19世纪30年代,德国科学家施莱登、施旺和魏尔肖等人创立细胞学说以来,细胞治疗的发展历程已经跨越了几百年的风雨。如今,它已广泛应用于骨髓移植、恶性肿瘤、自免、股骨头坏死、晚期肝硬化等疾病的治疗。
以CAR-T疗法为例,这一曾经针对艾滋病研究的技术,却在血液类癌症中大放异彩。
1989年,第一代CAR-T细胞技术崭露头角;
2017年,一纸批文首款CAR-T产品获批上市,标志着细胞治疗开启了新纪元;
迄今为止,国外已有6款CAR-T疗法产品获得FDA批准上市。
在全球生物医药的竞技场上,国际巨头虽凭借先发优势占据一席之地,但它的力量同样不容小觑。它以坚定的步伐,已然走在了生命科学的前沿,不断为人类健康事业贡献着中国智慧和中国力量。
01
临床研究
截至2024年8月,全球使用CAR-T疗法开展的肿瘤临床研究高达844项,中国以405项的份额占据了近半壁江山。
02
产品商业化
2021年
复星凯特和药明巨诺的CAR-T产品率先上市
2023年
驯鹿/信达和合源生物的CAR-T产品接踵而至
2024年
科济药业的CAR-T产品也加入了这一行列
03
国际合作
2017年
传奇生物与强生旗下的杨森制药签订了全球独家许可和合作协议,共同开发和商业化BCMA靶向的CAR-T疗法
2023年
阿斯利康宣布以约12亿美元的总价格收购细胞治疗企业亘喜生物
一个又一个跨国药企与新兴生物技术公司成功合作,标志着中国企业在部分新技术上已经处于全球领先水平,受到全球认可。
它,是中国生物行业。
后 记
这,是中国生物行业的故事,一个关于坚持与创新的故事。虽然道路漫长且充满挑战,但正如古语所言:道阻且长,行则将至。每一步的坚持,都是向着光明未来的迈进。尽管国产化的道路充满艰辛,早期产品或许尚存不足,但坚持是唯一的选择。毕竟,手中无剑和有剑不用是两回事。
编辑:芝麻核桃
声明:本文仅代表作者个人观点,不代表任何组织及本公众号立场,如有不当之处,敬请指正。如需转载,请注明作者及来源:蒲公英Biopharma。
“BLA杯”征文
《说说生物制药“国产化”那些事》
本次征文活动由“生物制药国产化推进联合会(BLA)”全程支持,围绕《说说生物制药“国产化”那些事》主题展开,可从政策法规、行业标准、研发技术、产业布局、设备耗材国产化、投融资、药品出海等多方面,谈谈你看到的、你所经历的、甚至你设想的生物制药“国产化”的那些事,期待您的投稿!
BLA杯征文合集
◆从政策、技术、出海等多维度探索生物制药国产化替代之路
◆不断向“新”,生物制药赛道国产化关键环节的“破壁者”
◆国产设备精益求精,无菌灌装系统“准、智、效”
◆国产洁净服飞速发展,为洁净防护领域注入新活力!
◆如何评估生物制品组件的相容性研究助力BLA申报?
◆看合成生物学技术如何助力重组胶原蛋白
◆中国药厂建设供应链随药企“出海”走向世界
◆如何才能使国产生物药走出去?
◆国产生物药的海外征途——创新与挑战并行
◆论生物制药国产化的未来
◆创新药研发之路
◆给国产生物药出海提建议
◆生物制药国产化之路到底为什么这么难?
◆关于CGT药物国产化研发的思考
◆浅谈生物制药关键工艺设备国产替代的变更考虑要素
关于BLA
生物制药国产化推进联合会(Biopharma Localization Association),简称BLA,总部位于上海,成立于2021年8月,是集生物制药领域平台、媒体、研发、生产、上下游供应链等企业的联合型社团组织。
BLA致力于助推生物制药“国产化”的三个核心:
▲ 助推建立健全中国生物药政策标准
▲ 助推研发制造安全有效的中国生物药
▲ 助推做大做强中国生物药国产供应链产品和体系
全面推进中国生物医药产业高质量发展,向生物科技强国和产业强国迈进。
BLA,for Biopharma,for China.
入会咨询:张老师,13062776509(微信注明:BLA)
邮箱:blachina@ouryao.com
疫苗生物类似药
2024-08-05
作者:严家新
目前丽沙病毒属的病毒,即(经典)狂犬病毒和狂犬病毒相关病毒,总共发现有大约18种,可细分为三个系统群(phylogroups)。现有狂犬病毒疫苗是用(经典)狂犬病毒(RABV)制备的,只能针对系统群1的病毒提供较好的免疫保护作用,而对系统群2和3的病毒则效果可能不佳。为了确保人类完全免除所有类型狂犬病毒的危害,有必要开发广谱狂犬病疫苗,能针对各种类型的狂犬病毒都发挥免疫保护作用。由于基因1型狂犬病毒(RABV)以外的17种丽沙病毒绝大多数都以蝙蝠为宿主,目前已知它们对人的危害很罕见,所以长期以来对此类疫苗研发的需求和动力不足,因而经费来源有限,进展缓慢。美国宾州费城的杰弗逊大学和美国CDC的研究人员曾合作在广谱狂犬病毒疫苗研究方面取得重要进展,他们的研究论文于2020年7月21日发表在《Cell Reports (细胞报告)》杂志上,本博客当年曾作过介绍(见相关博文)。
美国费城杰弗逊大学的研究人员在本月10日的《Viruses(病毒)》杂志上又发表论文介绍他们在此项研究中的最新进展。现将这篇新论文的主要内容进行摘译。
摘要:
除(经典)狂犬病毒(RABV)外,世界范围内还发现了大约16种类似狂犬病毒的丽沙病毒。已经描述了与经典狂犬病无关的人类死亡,但病例数量未知,而且这些丽沙病毒引起人类疾病的可能性是不可预测的。目前的狂犬病疫苗可能不能预防某些不同的丽沙病毒,如莫克拉(Mokola)病毒(MOKV)或拉各斯(Lagos)蝙蝠病毒(LBV)。因此,需要一种更广谱的丽沙病毒疫苗。在本文中,我们评估了一种新型的丽沙病毒疫苗,该疫苗使用了一种减毒的RABV载体,其中含有嵌合RABV糖蛋白(G),其中MOKV的抗原位点1取代了狂犬病毒的相应位点(RABVG-cAS1)。利用重组疫苗对小鼠进行免疫,并与同类疫苗进行免疫应答分析。我们的研究结果表明,疫苗RABVG- cAS1具有免疫原性,并诱导了针对RABV G和MOKV G的高滴度抗体。不同丽沙病毒的攻毒研究表明,用MOKV G的相应位点取代RABV G的单个抗原位点,比同源RABV疫苗有显著改善,并能同时预防RABV、伊尔库特病毒(IRKV)和MOKV。这种抗原表位嵌合的策略为构建广谱丽沙病毒疫苗铺平了道路,从而可安全地对抗由这些病毒引起的疾病。
1.前言
丽沙病毒属于丽沙病毒科的丽沙病毒属。除经典狂犬病毒(RABV)外,在非洲、亚洲、澳大利亚和欧洲还发现了来自狂犬病毒属的另外大约16种病毒。这些病毒在遗传上存在差异,具有不同的抗原性,可分为三个系统群:系统群I、系统群II和一个未分类的群[1 - 3]。尽管它们的基因和抗原性不同,但所有丽沙病毒都能在人类中引起致命疾病,症状与狂犬病相似。
狗仍然是人类暴露于经典RABV的主要原因,特别是在犬类疫苗接种计划没有有效执行的地区,如亚洲和非洲。据世界卫生组织(WHO)估计,每年约有5.9万人死于狂犬病,其中40%为15岁以下儿童[4]。
北美为根除狂犬病所做的密集努力取得了成功,导致每年的人类死亡人数减少[2]。然而,典型的狂犬病仍然存在于野生动物中,如浣熊(Procyon lotor)、臭鼬(Mephitis)、豺(Canis)、狐狸(Vulpes)和蝙蝠(Chiroptera),并且在南美洲发现了RABV的新谱系,扩大了储存宿主的名单。美国疾病预防控制中心最近报告称,2009年至2018年,在10例狂犬病病例中,有7例是死于蝙蝠介导的狂犬病[5]。
近年来,越来越多的丽沙病毒被发现可引起人类致命性脑炎,如2000年发现的澳大利亚蝙蝠丽沙病毒[6,7],2006年发现的南非杜文哈格(Duvenhage)病毒[8],2019年发现的俄罗斯IRKV病毒[9],2022年发现的欧洲蝙蝠1型丽沙病毒[10],以及2014年和2006年分别发现的南非莫克拉(Mokola)病毒和拉各斯( Lagos)蝙蝠病毒[11 - 13]。暴露于丽沙病毒后,病毒在暴露部位感染组织,然后通过周围神经系统逆行转运,最终到达中枢神经系统(CNS)并引起脑脊髓炎。在人类中,该疾病通常以非特异性症状(发烧、不适和头痛)开始,随后是焦虑和躁动。当病毒到达中枢神经系统时,它以一种称为离心扩散的现象在周围神经系统中向外扩散,通过神经到达唾液腺等组织,直到整个神经系统衰竭,导致瘫痪和快速死亡[14]。
目前获得许可的狂犬病疫苗在适当的情况下对传统的RABV有效。对于暴露前,在第0、7天和第21或28天接种疫苗,而暴露后的接种计划则在5剂或4剂之间变化[15,16]。狂犬病疫苗保护的相关性取决于靶向病毒表面糖蛋白(RABV G)的中和抗体水平[17]。
狂犬病毒独特的表面RABV G是中和抗体的唯一靶点,人们一直在研究其在融合前后的晶体结构,并确定其抗原位点I、II、III、IV和一个次要的抗原位点“a”[18 - 20]。抗原位点1 (AS1)是一个线性表位,可利用单克隆抗体(mAb) CR57结合RABVG的六个残基KLCGVL (226-231位)进行鉴定[20,21]。其他抗原位点是具有构象且不连续的。丽沙病毒糖蛋白具有57%-78%的相似性[19,22],影响了RABV接种后对非经典狂犬病毒的丽沙病毒的交叉保护。既往研究表明,狂犬病疫苗对中和效价在0.5国际单位[IU]以上的RABV具有保护作用,对其他具有不同中和抗体效价截断点的系统群1的丽沙病毒也具有交叉保护作用[20]。然而,目前的RABV疫苗对来自系统群2的丽沙病毒,如MOKV或属于未分类系统群的非常不同的丽沙病毒,不能诱导中和抗体,也不能提供保护作用[23-25]。
当前的狂犬病疫苗的这一局限性,以及非狂犬病毒丽沙病毒的不断出现[3,26],强调了存在尚未经评估的危险,以及需要一种能够预防由丽沙病毒属的多种病毒引起的人类狂犬病的更广谱有效的丽沙病毒疫苗。之前,我们的实验室设计了一种广谱丽沙病毒疫苗,其中RABV G和MOKV G之间的结构域被交换,产生了嵌合的RABV G,但是使用含有嵌合G的疫苗接种会损害对系统群1中其他丽沙病毒的保护,这导致我们试图重新设计疫苗,以更好地提供针对系统群I的保护[1]。利用结构生物学的最新进展和RABV G和MOKV G的结构[18,19,27],我们的论文提出了一种新的基于RABV G的疫苗,其中包含嵌合AS1的“RABVG- cAS1”,其中来自MOKV G的单个AS1被引入RABV G,取代了真实的AS1。
4.讨论
结构生物学及其相关工具的迅速发展,包括计算生物学、X射线晶体学和单克隆抗体图谱(mapping),已经解析了许多病毒的糖蛋白结构,导致了疫苗开发方法的革命[37]。在本文中,我们提出了一种针对狂犬病毒的下一代疫苗。特别关注狂犬病毒的AS1和莫科拉病毒糖蛋白,以设计和开发嵌合RABV-cAS1疫苗。RABV-cAS1针对系统群I和II狂犬病毒的多种病毒株具有保护作用,同时不会影响对RABV的免疫原性和保护能力。
我们证明,作为一个线性抗原位点[19,21],AS 1(222至252位)是两种不同丽沙病毒(RABV和MOKV)之间替代的功能选择,同时保持重组病毒稳定的复制。先前的研究也试图使用不同的方法开发广谱丽沙病毒疫苗,例如利用活载体表达丽沙病毒糖蛋白,利用多种疫苗结构,以及利用重组狂犬病毒表达丽沙病毒糖蛋白或单一嵌合糖蛋白[29,38]。使用重组狂犬病毒和重组痘苗病毒表达狂犬病毒糖蛋白的活载体策略可能因安全性问题而难以获准用于人体 [38, 39]。建议使用多种疫苗构建体,但这种策略对于低收入和中等收入国家来说成本较高 [40, 41]。最近,一种编码两种G蛋白(RABV G和MOKV G)的重组狂犬病毒导致病毒生长速度降低,并观察到G蛋白的不稳定性 [29]。因此,研究人员基于一种相关的弹状病毒(Rhabdovirus)—口蹄疫病毒在融合前和融合后的结构,创建了一种由MOKV G和RABV G组成的单个嵌合体G [42]。其中,携带嵌合体G的RABV之一(LyssaVax)表现稳定,而另一株则表现出低效的病毒传播和低感染性病毒滴度 [29]。
Fisher等人通过鼻内攻击测试了LyssaVax针对rRABV和rMOKV两种活病毒的保护能力。虽然这种途径是人工的,与自然的狂犬病感染途径不同,但它们对rRABV和rMOKV具有良好的保护作用[29]。同一组研究了LyssaVax对来自系统群I和II的一组LyssaVax病毒产生的血清学模式,但不包括系统群III。虽然LyssaVax可以刺激VNA(病毒中和抗体)滴度来对抗系统群II病毒,但研究表明,它在刺激VNA对抗系统群I中的非RABV病毒,尤其是IRKV和Duvenhage病毒(DUVV)的能力有所下降[29]。Fisher等人的研究确实表明,他们用于开发疫苗的“结构域(domain)”策略减少了一些针对系统群I中不同的丽沙病毒的VNAs的损失。通过在RABV- G载体疫苗中插入一个单一抗原侧面(MOKVG AS1),我们假设在保持对RABV的免疫原性的同时,扩展了对MOKV的免疫原性。本研究显示对MOKVG和RABVG均有较高的IgG滴度。同时,ELISA法对狂犬病免疫小鼠血清观察到异种抗原之间存在交叉反应,与既往研究所显示的相同[29];这些抗体并未介导VNAs(病毒中和抗体)。
我们还假设,与标准疫苗相比,嵌合抗原位点(222-252位)可以提供更广谱的保护,抵御各种丽沙病毒的攻击,同时仍然保持类似的针对经典狂犬病的保护。RABV-cAS1显示了MOKV的AS1在维持针对系统群I的VNA的同时引发针对系统群II病毒的中和抗体的潜力。
这些结果与其他研究一致,这些研究表明每个抗原位点在产生中和抗体时具有特异性。我们的研究结果证实了基于AS1的疫苗方法能够产生持续的免疫应答和特异性中和抗体[43-45]。
在我们的研究中,我们证明了RABV- cAS1 能够产生针对系统群I中两种不同病毒(RABV和IRKV)的中和抗体,而且没有显著差异。通过我们的新方法,这似乎是对嵌合LyssaVax疫苗的重大改进,因为Fisher等人先前发表的尝试表明,在有佐剂和没有佐剂的情况下,LyssaVax丧失了针对IRKV产生VNA的刺激作用[26]。这可能与用于创建嵌合G的设计有关,其中RABVG的抗原位点的更多抗原部分被牺牲。值得注意的是,参与保护的抗原位点并不一定对所有的丽沙病毒都相同。RABV疫苗在系统群I内可出现交叉保护;然而,保护水平是不明确的,可能取决于高水平的 G特异性抗体[46 - 48]。我们的发现与其他研究结果一致,表明现有狂犬病疫苗对IRKV的保护作用最小[48,49]。这种有限的保护最可能的解释是系统群I丽沙病毒内部的抗原变异,因为IRKV与欧洲蝙蝠丽沙病毒和杜文海格(Duvenhage)病毒的关系比与狂犬病毒(RABV)的关系更密切[1,50]。RABV与IRKV的比较表明,抗原位点 I、II和 III 不同,而抗原位点 IV 则完全保守[49,51]。
虽然本文的交叉保护评价仅限于IRKV,但与其他蝙蝠相关的丽沙病毒相比,该丽沙病毒在各种研究中显示出最高的致病性,这解释了为什么现有的基于 RABV的生物制剂对IRKV的攻击只具有部分保护作用[49,51]。此外,在祖先水平上,IRKV、RABV、ABLV、ARAV、BBLV、KHUV、DUVV、EBLV-1和EBLV-2病毒都被归类为一组(系统群I),这表明这些病毒之间应该存在血清学交叉反应[24,50]。其他研究将系统群I分为两个主要世系(lineages):一个包括IRKV、EBLV-1和DUVV,另一个世系包括古北区(Palearctic)(ARAV、BBLV、KHUV和EBLV-2)、澳大利亚(ABLV)、东方(GBLV)和美洲(RABV)丽沙病毒物种[52]。
有趣的是,用RABV-cAS1疫苗免疫的小鼠对wt(野生型)IRKV具有完全的保护作用,而用RABV疫苗免疫的小鼠的保护作用有限。一方面,AS1含有一个具有关键残基LCGV的线性表位,负责CR57和62071-3 mAbs的结合。IRKV AS1与RABV AS1的比对显示IRKV丢失了V230。这种氨基酸变化可能改变单克隆抗体在该线性表位内的结合,扰乱中和作用并限制针对IRKV的保护。最近的研究支持了我们的发现,表明RABV疫苗在动物模型中对IRKV并不完全有效[49,51]。与RABV相比,AS1如何提高针对IRKV的保护尚不清楚。RABV、IRKV和MOKV AS1的计算机分析表明,IRKV AS1与RABV AS1的同源性为64.5%,而与MOKV AS1的同源性为61.3%。然而,IRKV AS1有两个残基R245和D247与MOKV AS1共享,而与RABV-cAS1不共享(补充图S3)。为了证实这一点,未来对这两个残基的分析可能会更好地理解通过MOKV AS1获得的保护。
我们的研究结果强调,虽然MOKV免疫的小鼠得到了充分的保护,但RABV-cAS1疫苗可提供40%的保护,比狂犬病疫苗有显著提高,而狂犬病疫苗对MOKV没有保护作用。这些发现与先前的研究一致,证实RABV疫苗不能针对系统群II的丽沙病毒提供保护[24,25,53]。虽然对wt MOKV的保护只是部分的,但我们认为这与疫苗接种程序无关。需要强调的是,RABV-cASI疫苗的灭活和佐剂添加,采用的是5µg的TLR-4激动剂合成的单磷酰脂质A (Monophosphoryl Lipid A,MPLA),3D (6A)-PHAD (PHAD),制备成含2%角鲨烯(squalene)的油乳剂(SE)。最近对狂犬病载体疫苗的研究表明,使用PHAD-SE作佐剂的灭活疫苗明显改善了疫苗引起的免疫应答[32-34]。
此外,我们在死于疾病的小鼠死后脑组织中发现了MOKVG,但在存活的小鼠中却没有。目前还不清楚疫苗是否能够阻止病毒进入大脑,或者在病毒进入大脑后将其清除。攻击后症状数据显示,经RABV-cAS1免疫的小鼠和经鼻注射wtMOKV的小鼠出现毛发皱褶、嗜睡和体重减轻,但未观察到瘫痪。当对照组在第9天至第10天被安乐死时,两只幸存的老鼠不符合安乐死标准,并在第11天开始恢复。根据图8K所示脑组织的RT-PCR结果,在同一组的三只安乐死小鼠的脑组织中发现了wt MOKV病毒,而在这两只幸存者的脑组织中没有发现。因此,我们推测病毒到达大脑,然后小鼠仍然可能康复,特别是因为wt MOKV攻击是在鼻内进行的。这条途径被证明是快速的,并确保通过嗅觉和三叉神经通路渗透大脑而感染[54-56]。
狂犬病感染后康复是可能的,在人类临床感染和小鼠、狗、雪貂和兔子等动物的实验感染中都有报道[57 - 59]。一旦病毒进入CNS(中枢神经系统),它就会脱离体液免疫反应的保护,除非抗体能够穿过BBB(血脑屏障)。既往研究发现康复与BBB通透性和抗体在脑脊液中的存在相关[59 - 61]。根据已发表的数据,T细胞的活性可以抑制狂犬病毒的复制[62,63],但通过B细胞浸润穿过BBB而产生狂犬病毒特异性抗体对于消除狂犬病毒至关重要[17,61,64]。在正常情况下,BBB阻止血液中的细胞和因子与CNS中的细胞和因子之间的接触。在从CNS组织清除减毒狂犬病毒的过程中,BBB向液相标记物开放,但不向大分子开放。这可能允许神经血管系统中的免疫细胞检测在CNS中产生的化学引诱剂(chemoattractants),并沿着BBB的梯度向上移动到感染组织[61,64]。这些发现与我们的数据一致:狂犬病感染可能在病毒到达CNS组织后仍然得到恢复。
我们的研究提出了一种开发广谱丽沙病毒疫苗的新方法。然而,这项研究有一些局限性。由于MOKV不通过肌肉内途径致病,因此MOKV攻击是通过鼻内途径进行的,这是一种人工途径,由于其靠近CNS,增加了导致严重疾病的风险。此外,由于血清数量有限,我们知道能用于进行交叉保护试验的非狂犬病丽沙病毒的数量有限,而我们认为阈值的确定才能有助于更深入的了解。针对系统群II的其他丽沙病毒拉,例如拉各斯蝙蝠病毒和希莫尼(Shimoni)蝙蝠病毒的保护研究,可能会揭示使用相同的攻击途径具有更好的保护作用,从而能更好地进行数据比较。
重要的是,尽管这种方法新颖,但对系统群I和II的完全保护尚未实现,而且必须扩展到系统群III。开发广谱丽沙病毒需要一种新的策略,在生产稳定的重组病毒和具有丽沙病毒属代表的更广谱抗原的嵌合单糖蛋白之间取得平衡。
5.结论
由于迫切需要一种广谱丽沙病毒疫苗,本研究提出了一种创新的广谱丽沙病毒疫苗策略,将RABVG的抗原位点1替换为MOKVG的抗原位点1。在保持嵌合RABV- G的功能及其对RABV的保护能力的同时,嵌合疫苗能够提供针对IRKV(系统群I中的一种远缘丽沙病毒)的完全保护,以及对MOKV(系统群II中的一种远缘丽沙病毒)的部分保护。
我们的研究是朝着有效的广谱丽沙病毒疫苗迈出的第一步,该疫苗可预防由多种丽沙病毒引起的狂犬病等疾病,目前尚无疫苗或其他对策可供使用。
参考文献:
Ben Hamed, S.; et al., Toward the Development of a Pan-Lyssavirus Vaccine. Viruses 2024, 16, 1107.
为了推动人用及兽用疫苗行业交流,共同探讨该领域的最新研发进展、产业化现状及未来发展趋势,生物制品圈联合四叶草会展将于2024年8月16日-17日在苏州共同举办“第三届新型疫苗研发峰会”。诚邀全国相关领域专家、学者和企业家共享学术盛会。现将有关事宜通知如下。
名称:2024第三届新型疫苗研发峰会
时间:2024年8月16-17日(周五-周六)
地点:苏州(酒店定向通知)
主办单位:四叶草会展、生物制品圈
媒体支持:药时空、抗体圈、细胞基因研究圈
会议费用:点击二维码即可查询。
报名方式:扫描下方二维码→ 填写表格 → 报名成功(报名志愿者,免交报名费,承担一定工作任务)!
转发分享福利:疫苗研发企业、科研院校和监管机构人员,前30位报名人员转发分享会议信息可免费参会(不含餐饮)报名后,扫描下方二维码加工作人员微信后,发送截图即可。
组委会获得报名信息后,根据报名信息进行初筛,并进一步与报名者沟通确认,实现精准邀请,最终有机会进入大会微信群(严格审核通过)。
大会日程
已经确定的报告人
识别微信二维码,添加生物制品圈小编,符合条件者即可加入
生物制品微信群!
请注明:姓名+研究方向!
版
权
声
明
本公众号所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,不希望被转载的媒体或个人可与我们联系(cbplib@163.com),我们将立即进行删除处理。所有文章仅代表作者观点,不代表本站立场。
疫苗
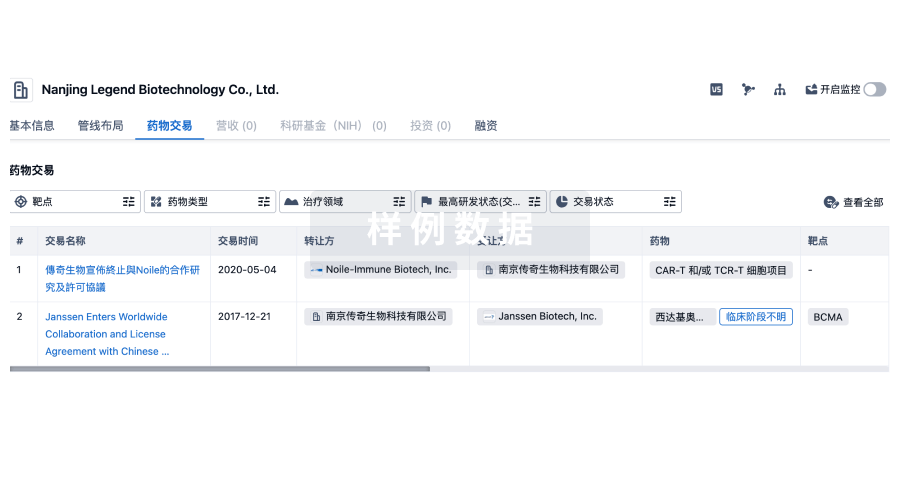
100 项与 浙江东方绍兴酒有限公司 相关的药物交易
登录后查看更多信息
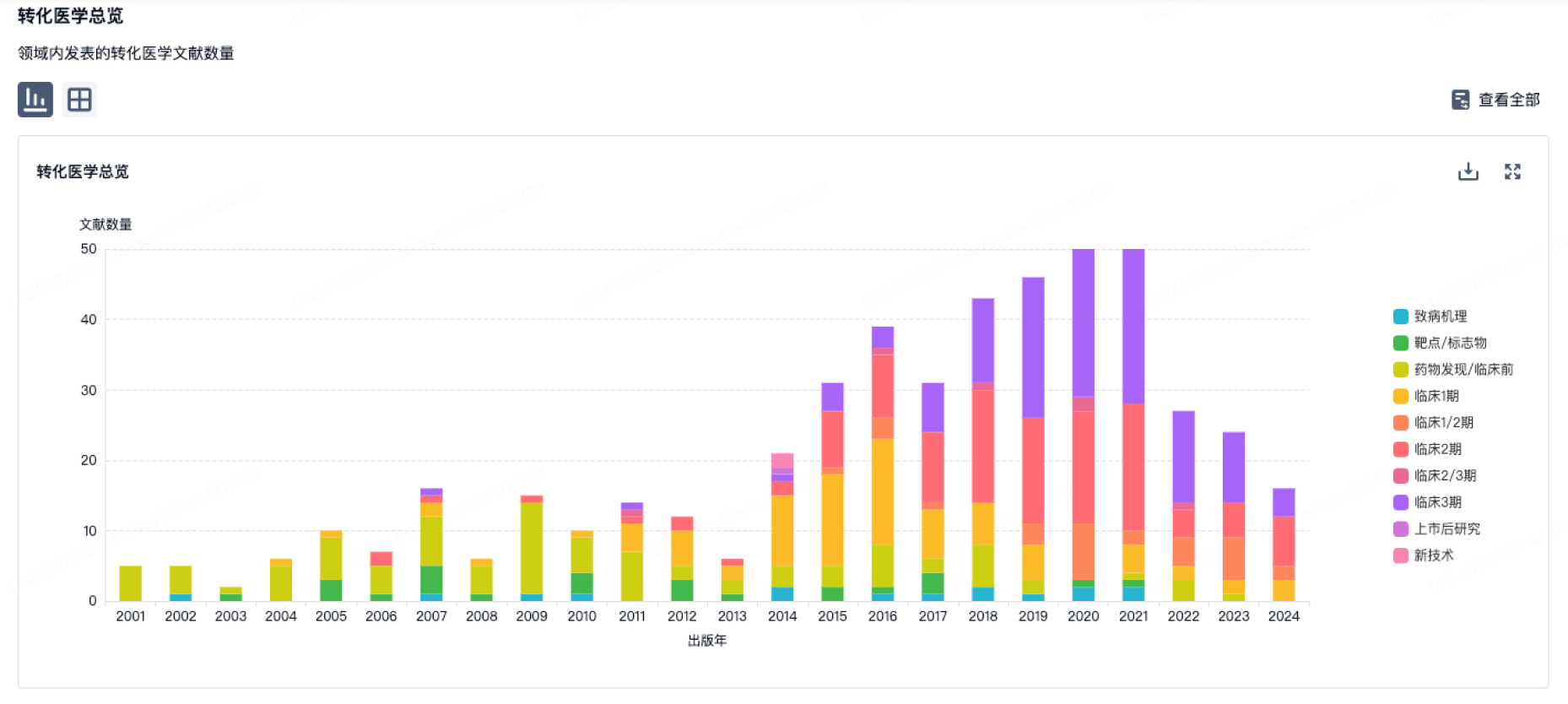
100 项与 浙江东方绍兴酒有限公司 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2024年10月06日管线快照
无数据报导
登录后保持更新
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
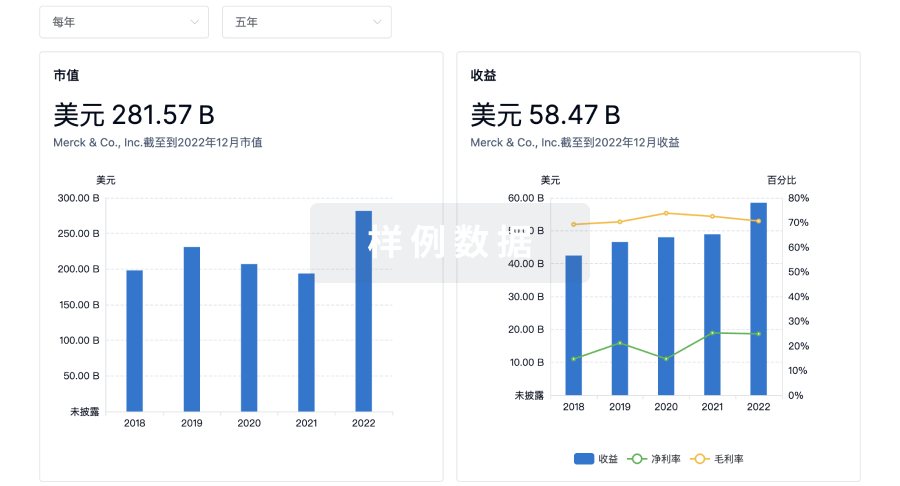
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
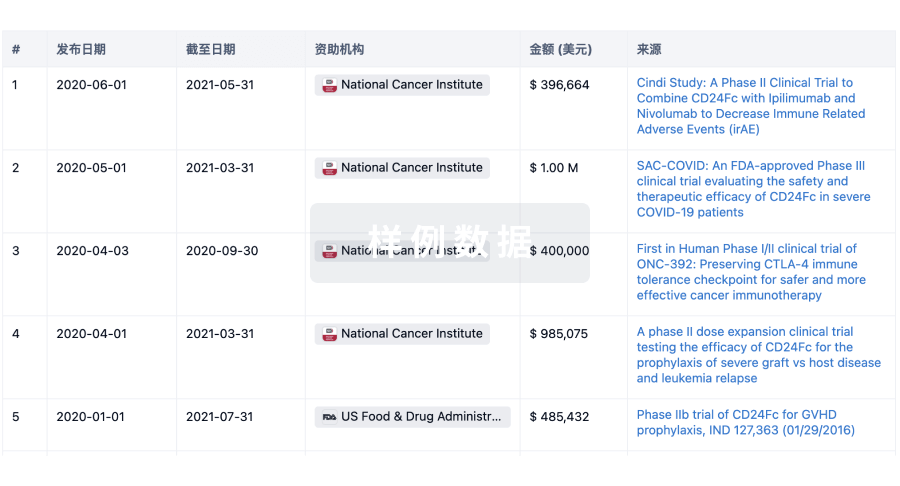
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
标准版
¥16800
元/账号/年
新药情报库 | 省钱又好用!
立即使用
来和芽仔聊天吧
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用