预约演示
更新于:2025-11-01
V-0219
更新于:2025-11-01
概要
基本信息
非在研机构- |
权益机构- |
最高研发阶段临床前 |
首次获批日期- |
最高研发阶段(中国)- |
特殊审评- |
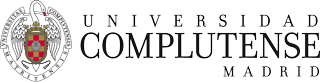
结构/序列
分子式C20H25F3N4O2 |
InChIKeyVFQGZIAZHIRPPQ-UHFFFAOYSA-N |
CAS号878453-71-5 |
关联
100 项与 V-0219 相关的临床结果
登录后查看更多信息
100 项与 V-0219 相关的转化医学
登录后查看更多信息
100 项与 V-0219 相关的专利(医药)
登录后查看更多信息
1
项与 V-0219 相关的文献(医药)2022-04-14·Journal of medicinal chemistry1区 · 医学
Discovery of V-0219: A Small-Molecule Positive Allosteric Modulator of the Glucagon-Like Peptide-1 Receptor toward Oral Treatment for “Diabesity”
1区 · 医学
Article
作者: Suárez, Juan ; Lutz, Beat ; Garcia-Ladona, F. Javier ; Alén, Francisco ; Rodríguez de Fonseca, Fernando ; Rodríguez, Ramón ; Brea, José ; Orio, Laura ; Alonso, Mónica ; Cruces, Jacobo ; López-Rodríguez, María L. ; Ballesteros, Juan A. ; Benhamú, Bellinda ; Baixeras, Elena ; Bennett, Teresa A. ; Srivastava, Raj Kamal ; Decara, Juan M. ; Codesido, Eva ; Vázquez-Villa, Henar ; Loza, María I. ; García-Cárceles, Javier ; Escobar-Peña, Andrea
Peptidic agonists of the glucagon-like peptide-1 receptor (GLP-1R) have gained a prominent role in the therapy of type-2 diabetes and are being considered for reducing food intake in obesity. Potential advantages of small molecules acting as positive allosteric modulators (PAMs) of GLP-1R, including oral administration and reduced unwanted effects, could improve the utility of this class of drugs. Here, we describe the discovery of compound 9 (4-{[1-({3-[4-(trifluoromethyl)phenyl]-1,2,4-oxadiazol-5-yl}methyl)piperidin-3-yl]methyl}morpholine, V-0219) that exhibits enhanced efficacy of GLP-1R stimulation, subnanomolar potency in the potentiation of insulin secretion, and no significant off-target activities. The identified GLP-1R PAM shows a remarkable in vivo activity, reducing food intake and improving glucose handling in normal and diabetic rodents. Enantioselective synthesis revealed oral efficacy for (S)-9 in animal models. Compound 9 behavior bolsters the interest of a small-molecule PAM of GLP-1R as a promising therapeutic approach for the increasingly prevalent obesity-associated diabetes.
1
项与 V-0219 相关的新闻(医药)2024-08-26
摘要:G蛋白偶联受体(GPCRs)是最大且最多样化的细胞表面受体家族,具有广泛的配体和功能。自从1986年第一个GPCR被克隆和表达以来,我们对这个受体类别的药物发现已经学到了大量的知识,以至于现在GPCRs作为批准药物的最成功靶标类别已经得到了广泛认可。在这里,我们带领读者经历从靶标到临床的GPCR药物发现之旅,强调关键的学习点、最佳实践、挑战、趋势和洞察,这些对于发现最终在患者中调节GPCR功能以治疗的药物至关重要。GPCR药物发现的未来是鼓舞人心的,更多理想的药物机制和新技术正在使交付更好、更成功的药物成为可能。
1.引言
GPCRs是发现新药最成功的靶标之一,代表着由约821个人类基因编码的细胞表面跨膜受体的最大超家族;见的综述。实际上,据估计约34%的上市药物是GPCR功能的调节剂,而大约140个GPCR的天然配体和功能尚未被发现——所谓的孤儿GPCRs。这篇综述集中在小分子GPCR药物发现上,因为大多数GPCR药物是小分子(约92%)而不是像抗体这样的生物制剂。这种药物模式的当前偏好主要是由于历史上GPCR具有小分子天然配体的强烈记录,加上生成受体特异性生物药物的技术难度。凭借这一广泛的功能范围,GPCR药物几乎出现在所有治疗领域,特别是神经学、心血管和炎症。有趣的是,尽管通过针对免疫细胞(例如,在免疫肿瘤学中的腺苷A2AR)的GPCR药物在肿瘤学中的存在感较少,但随着新ADCs的出现和新的疾病联系,这种状况将会增加。许多专家认为,60-85%的潜在治疗性GPCR尚未被药物化。值得注意的是,对于一些批准的GPCR药物,疾病疗效是由受体多药理学驱动的,无论是设计上的(例如,用于2型糖尿病的共激动剂肽Tirzepatide)还是偶然的(例如,非典型抗精神病药Clozapine)。获得可接受的治疗窗口的正确多药理学量是一个挑战,并且在项目开始时很难预测。所有GPCRs都表现出多功能的七跨膜框架,能够容纳各种大小的配体激活受体,从小质子到大蛋白质以及不同的物理形态,从光子到蛋白酶激活。此外,GPCRs的激活是通过许多综合生物学过程介导的,并与多种细胞功能相关联。有了这么多不同的配体和药理学,GPCRs表现出许多生理和病理生理作用,因此它们继续吸引大量的药物发现和科学兴趣。人类GPCRs被分为六个主要家族(A:视紫红质,B:分泌素,C:谷氨酸,D:真菌,E:cAMP和F:Frizzled),其中视紫红质(Class A)家族是最被研究的(94%的GPCR药物)。大多数GPCR药物要么是拮抗剂要么是激动剂(分别为53%和42%),但越来越多有利的、精细的模式,具有改进的GPCR选择性,如负性和正性异构调节剂(NAM和PAMs)和偏置配体在临床上越来越受到关注。这篇综述将突出从靶标验证到临床的现代GPCR药物发现的关键方面(见图1)。每个方面都将由相关的案例研究、挑战和最新的知识支持,然后是对这一激动人心的药物发现领域的未来展望的总结性思考。在发现阶段开始时,候选GPCR药物靶标应经过严格的评估,以确定药物靶标的可取性、发现项目的可行性和投资。这个过程涉及一系列考虑因素。关键要素包括靶标验证、疾病联系、靶标药物性、检测可行性、多靶标选择性和在进一步投资新药物发现项目之前的命中发现策略。图2展示了一个通用的命中发现过程,说明了所有当前可用于识别小分子化学起点的方法,但对于GPCRs来说,重要的途径是利用现有药理学的基于知识的和通过HTS的多样性筛选。尽管GPCR结构和生物物理信息正在增加并且非常理想,但通常更难实现并前瞻性地应用于药物发现。
图1. GPCR药物发现 - 从目标到临床的流程。
图2. 通用的候选药物发现流程。
在介绍了一般的药物发现过程和GPCR特定的命中发现方法之后,我们将进入更详细的部分,涵盖HTS、定义药物功能的检测、机制和驱动DMTA(设计-制造-测试-分析)周期的检测,GPCR蛋白表达,生物物理学,GPCR结构信息的洞察,作为进入药物和计算化学的过渡,最后是关于新兴GPCR药物的后期转化生物标志物和临床方面的部分。总之,这篇综述将探讨GPCR药物发现科学和疾病理解如何预测发展,推动新靶标、概念和有价值的药物模式,以识别更好的药物治疗。GPCR药物发现继续是一个生产力很高的新药领域(例如,2021年批准的首创C5a受体拮抗剂Avacopan用于治疗血管炎)。随着最近的趋势,如用于治疗抑郁症的迷幻剂和新的模式如偏置激动剂,这一靶标类别的新药物发现的未来看起来很有希望。
1.1.HTSHTS是一种用于快速识别药物发现活动起始分子的方法,在这种方法中,成千上万的化合物被筛选针对一个目标。一个成功的HTS活动需要显著的基础设施,包括一个高质量的结构多样化小分子库和多个科学学科的整合,包括药物化学、计算化学、信息学和生物学。HTS检测必须经过良好的验证,以确保它们高度可重复、稳健、低成本,并与检测自动化兼容。GPCRs历来是HTS活动最受欢迎的目标之一。对GPCR HTS的需求随时间波动;然而,近年来,GPCR HTS数量增加的趋势显著。GPCR HTS活动最常用的方法涉及研究受体激活下游的细胞内事件。例如,这些方法测量第二信使,如Ca2+、cAMP(环磷酸腺苷)、pERK(磷酸化细胞外信号调节激酶)和IP1(肌醇一磷酸)、基因表达报告基因,以及β-arrestin对GPCR的招募。
现在有许多HTS兼容的检测技术可用于测量这些GPCR第二信使。HTRF(均质时间分辨荧光)和AlphaScreen非常适合HTS,以均质格式产生一致的高质量数据,信号和试剂稳定性令人印象深刻,与检测自动化兼容,并且可以扩展到384和1536孔格式。一个很好的例子是Myers等人在2020年使用的‘cAMP Gs动态试剂盒HTRF®’检测,用于识别 apelin 受体的小分子激动剂。有许多其他方法可用于测量GPCR第二信使,包括最近用于从超过300,000种化合物筛选中识别35种选择性、反向激动剂GPR3的GloSensor™ cAMP检测。
测量细胞内钙瞬变是HTS Gαq偶联GPCRs的常用读数。FLIPR系统(Molecular Devices)和Hamamatsu Photonics FDSS仪器是细胞内钙测量的行业领先技术。这些技术利用敏感的钙染料来测量细胞内钙的变化。还有许多其他基于钙的筛选方法可用,包括最近开发的适用于CCR6受体的HTS兼容钙动员检测,该检测使用己糖胺酶活性作为荧光读数。
有几种HTS兼容的方法可用于测量β-arrestin GPCRs的招募,但需要修改GPCR C-末端,这可能会干扰受体的活性和信号。在2021年,Hauge Pedersen等人开发了一种名为‘MeNArC’的基于发光的β-arrestin招募检测,不涉及修改GPCR。该方法利用了一种新型NanoLuc分裂,其中N-末端NanoLuc片段‘MeN’锚定在膜上,C-末端NanoLuc片段‘ArC’与β-arrestin融合。这种方法尚未用于HTS,然而,初步数据表明这种方法将适合于未来GPCRs的HTS。
无细胞筛选纯化的GPCRs具有减少非靶标效应的优势。然而,以足够的规模为HTS生产纯化的重组膜蛋白在生物学上相关的构象已被证明具有挑战性,并限制了在无细胞环境中筛选GPCRs的能力。尽管如此,有几种无细胞HTS兼容的筛选方法可用,例如用于筛选神经肽受体1型(NTS1)配体的无细胞荧光偏振检测。质谱是一种常用的技术,用于识别蛋白质-配体相互作用,并已成功用于近年来的许多GPCR HTS活动中。在2019年,Lu等人使用亲和质谱筛选识别了A2A腺苷受体的三种新拮抗剂。随后,他们在2021年使用类似的方法识别了腺苷A2A受体的NAM。
在某些情况下,传统的高通量筛选(HTS)化合物库已被DNA编码的化合物库(DELs)所取代,因为它们允许并行亲和筛选更多的小分子。DELs非常庞大,包含超过10^8个小分子,所有这些小分子都共价连接到一个独特的DNA条形码上。DNA标签充当条形码,可以通过读取来确定每个击中目标的小分子的身份。GPCR与DEL库一起孵育,通过亲和筛选鉴定与受体结合的化合物,然后放大并测序DNA标签。最初,DEL筛选仅限于可溶性蛋白靶标,但最近,DEL筛选已适应于膜受体,并于2015年首次被Wu等人用于筛选GPCRs,以鉴定NK3 tachykinin受体的拮抗剂,并为未来的GPCR HTS提供了一个有希望的方法。
除了筛选GPCRs的小分子调节剂外,HTS还可以筛选GPCR生物制剂,如肽和寡核苷酸。可以使用一种称为SELEX(通过指数富集的配体顺序进化)的方法筛选GPCR的核酸适配体,该方法涉及筛选数十亿个寡核苷酸对目标受体。在2021年,Takahashi等人使用基于SELEX的筛选方法鉴定了P2RY2 GPCR的正性异构调节剂。
1.2.GPCR筛选和筛选级联
一个综合药物筛选级联的目标是量身定制以满足总体项目目标(例如,发现一种口服GPCR拮抗剂)和项目阶段(例如,HTS后早期命中验证,见图3或支持后期阶段的主导优化DMTA循环)。级联顶部的主筛选(图3)旨在快速且稳健地测量化合物以期望的机制(例如,激动剂或拮抗剂)与GPCR结合的能力,通常在HTS中开始。有许多类型的GPCR检测,但它们通常分为两类,细胞内信号终点(如HTS部分所讨论)或受体结合检测。使用放射性配体、荧光偏振或GTPγS交换技术的GPCR结合检测,历来是识别GPCR命中和直接化学的最简单和最快的方法。这些检测具有优势以及挑战和限制。例如,结合检测非常擅长提供有关药物-靶标相互作用的信息,包括配体结合的动力学和目标停留时间。然而,除非化合物结合到正性配体结合位点,否则仅用结合检测很难推断有关信号通路或功能性效力的任何信息,因此在命中识别过程的早期结合受体结合和信号检测很重要。此外,当通过测量细胞内第二信使来评估化合物的效力时,必须使用内源性细胞或疾病相关组织模型确认作用机制(MOA)和所测量的响应将预测体内所需的治疗效果。大多数基于细胞的检测使用转染了目标GPCR的HEK293或CHO细胞系。在这种人为增强的系统中,由于“受体储备”的丰富性,目标受体的分数占据可能会高估激动剂的效力和效力。
图3. 典型的GPCR高通量筛选(HTS)拮抗剂候选药物验证生物学筛选级联。
通常与主检测并行,会运行一个反筛选以排除由于检测技术干扰或通过非靶标活性在主检测中识别的不需要的假阳性命中。通常,这些反筛选测量在主筛选中使用的相同信号通路和检测技术在缺乏目标受体的情况下击中活性的细胞背景。被识别为Pan-Assay Interference compounds (PAINs)的各种化学物质经常被重复识别为假阳性。这些化合物通常显示出固有的氧化还原活性,作为分子聚集体,或可能含有活性杂质,如重金属,或具有内在的荧光猝灭特性,干扰各种筛选技术。从筛选库中识别和排除PAINs通常具有挑战性,导致在主筛选后回顾性地识别假阳性命中。PAINs表现出非典型的药理学特征,如暗示超化学计量相互作用的陡浓度-反应曲线,代表聚集行为的洗涤剂敏感性,非竞争性和/或时间依赖性,以及化学系列中平坦的结构-活性关系(SAR)。理想情况下,反筛选后会进行一个正交检测,使用与主细胞信号筛选不同的筛选技术(例如,生物物理或生化检测),以确保活性不局限于特定的检测技术,并验证化合物与目标受体的结合。大多数GPCR药物发现活动中的另一个关键问题是早期测量化合物对相关、不需要的GPCRs的选择性,因为缺乏选择性可能需要在项目中解决。通常,选择性检测被内置到筛选级联的早期,以防止非选择性命中的进展,并允许药物化学家提高选择性。在物种受体同源物上使用人等效化合物的效力和机制是理想的,以便可以评估下游优化化合物的体内药代动力学和药效学属性。因此,最初使用人等效的重组GPCR物种检测来评估命中同源GPCR药理学。此外,在内源性表达细胞或组织检测中早期测试命中化合物也弥合了在过度表达的重组目标系统中观察到的GPCR效力和效力与在主要细胞靶标(例如,中性粒细胞中的CXCR1)之间的差距。当人和动物同源物之间的序列差异更大和/或药物通过较少保守的别构位点工作时,药物功能和/或效力的物种交叉较差的情况更频繁发生,见GPR35药理学作为一个例子,导致开发转基因敲入人GPCR小鼠进行PK/PD研究的额外费用。
我们现在更仔细地看看在药物发现筛选级联中使用的不同类型的GPCR检测,包括生物物理、生化结合检测、细胞内信号、表型和MOA研究。
1.3.GPCR表达和纯化
包括GPCRs在内的膜蛋白由于其复杂的结构、低表达水平和较差的稳定性而难以在高产量下产生和分离。GPCR表达的一个主要挑战是实现足够高的表达水平,以实现结构研究或功能检测等下游应用。低表达水平通常归因于GPCRs的折叠和插入到细胞膜中,这是一个复杂的过程。此外,在某些情况下,必须存在正确的翻译后修饰。此外,GPCRs可能非常不稳定,导致展开或降解,使得难以获得用于下游应用的纯净和稳定的蛋白质样品。下面探讨了克服这些挑战的策略和技术。
1.4.GPCRs过表达的细胞系统
人类GPCRs的重组表达是生产高产量孤立受体的重要先驱步骤,使功能、生物物理和结构表征检测能够发现针对这些受体的药物。GPCRs必须在能够适当处理其特定的翻译后处理和修饰的系统中表达,包括折叠和插入到膜中以及侧链的糖基化。表达系统的选择可以显著影响所产生蛋白质的质量和产量,以及生产所需的时间和成本。
一些GPCRs已成功在大肠杆菌中表达;然而,大肠杆菌是一个在成功表达GPCR方面的非常有限的系统,因此,这些例子往往是例外或需要对蛋白质序列进行广泛的修改才能兼容。同样,也测试了无细胞表达系统用于生产GPCRs。有一些成功的无细胞表达GPCRs的测试案例;然而,这项技术通常比成功生产GPCR的其他系统更有限制。因此,不推荐在大肠杆菌或无细胞系统中表达GPCRs。人类胚胎肾(HEK)细胞系统更广泛地用于过度表达GPCRs。该系统为表达人类GPCRs提供了一个类似的环境,并允许进行功能性GPCR表达所需的适当翻译后修饰。然而,使用HEK细胞可能具有挑战性,因为使用这些细胞的成本高且复杂。
首选的GPCR过度表达方法是使用昆虫细胞系统。这种偏好从提交给蛋白质数据银行(PDB)的GPCR结构数量中可以明显看出,这些结构在Hi5、Sf9或Sf21昆虫细胞系统中表达,占总结构的三分之二。昆虫细胞是生产GPCRs的合适宿主,因为它们可以执行复杂的翻译后修饰,包括糖基化,并促进受体的正确折叠和插入膜中。与其他表达系统相比,昆虫细胞具有几个优点。它们易于大量培养,并且可以产生高产量的重组蛋白。此外,它们比哺乳动物细胞系统更便宜、更不复杂,使它们成为大规模生产GPCRs的一个有吸引力的选择。
1.5.GPCR生产的构建设计和纯化策略
设计GPCR构建用于重组表达时需要考虑各种因素。纯化策略对于确定必须添加到受体的标签很重要。此外,特定的分析技术可能需要额外的标签和融合。此外,受体的下游使用决定了构建的设计。例如,用于表面等离子共振(SPR)实验的GPCR构建的要求可能与用于X射线晶体学或冷冻电镜实验的要求不同。最后,在使用最天然的受体序列和允许蛋白质以给定应用所需的规模、质量和稳定性产生的修改之间必须进行权衡。这些修改可能包括标签、截断、删除、融合伙伴插入和稳定突变。
为了纯化GPCRs,通常需要一个纯化标签。此外,由于GPCRs的低表达产量和纯化步骤中使用的高洗涤剂浓度,高度特异性的标签比常用的亲和标签(如6xHis标签)更有利。使用组氨酸标签至少需要8xHis标签,以提供对金属亲和树脂的更多特异性。或者,通常使用FLAG标签和双链strep标签,提供更大的树脂结合特异性,从而在纯化过程中的初始捕获步骤中得到更纯净的产品。抗体偶联的树脂提供了最高的树脂结合特异性,但价格昂贵且树脂结合能力有限。因此,在选择合适的纯化标签时,必须考虑树脂结合特异性和容量之间的权衡。用于膜蛋白,特别是GPCRs的一种标签是将构建物的天然C末端替换为来自牛视紫红质C末端的1D4表位。由于这个标签源自膜蛋白的两亲螺旋,因此添加这个标签对天然表达谱的影响比人工纯化标签更低。
除了纯化标签外,还可以添加额外的序列以提高GPCR的表达和检测。例如,在GPCR序列的N末端添加可切割的流感血凝素信号序列可以帮助增加GPCR的表达水平,并为构建物的纯化或检测提供另一种选择。提高GPCR构建物检测的另一种方法是在构建物中添加绿色荧光蛋白(GFP)融合,允许直接从细胞使用荧光信号快速评估GPCR的表达。添加GFP标签还通过使用偶联到树脂的抗GFP抗体提供纯化选项。对于使用SPR(见生物物理学部分)的生物物理受体-配体研究,常见的添加标签是avi标签,这将允许使用细菌BirA酶对蛋白质进行位点特异性生物素化。如果蛋白质与BirA酶共表达或在准备GPCRs进行SPR研究期间使用纯化的BirA进行体外使用,这种标记方法将起作用。
在成功纯化后,从受体中去除额外的纯化标签是标准做法,通过在受体和纯化标签之间添加蛋白酶切割位点来实现。常用的位点包括TEV蛋白酶的ENLYFQ|(S/G)、HRV蛋白酶的LEVLFQ|GP和Factor Xa蛋白酶的IEGR|,其中“|”表示切割位点,括号中的氨基酸表示不分等位位置。这些蛋白酶具有高度特异性和效率,使它们成为流行的选择。
GPCRs往往不稳定,这可能是纯化和结构确定的主要障碍。受体一旦从天然双层环境中移除,就会迅速展开或失去活性。为了应对使用GPCRs的这一挑战性方面,通常采用的方法是引入热稳定的点突变。虽然向GPCR序列中添加突变可能不理想,但有时为了便于使用受体是必要的。通常包括功能测试,以确保所选的突变稳定,但不干扰配体结合。这些突变可能通过减少构象灵活性来稳定,将GPCR锁定在单一状态,如活性或非活性状态。将受体锁定在单一状态对于通过X射线晶体学确定高分辨率结构很重要。通过X射线晶体学解决的所有GPCR结构中有三分之二至少经过了一种稳定突变的工程改造。
已经使用了多种方法来确定膜蛋白的稳定突变。扫描突变无疑是确定稳定突变的最成功方法,其中GPCR的每个位置都突变为丙氨酸(或另一种氨基酸替代物),并使用热熔曲线测试每个突变体的稳定性。这个过程通常是迭代进行的,构建可以增加GPCR稳定性的突变组合。或者,定向进化通过随机突变也显示出在确定这些稳定突变方面的成功,其中实验负担降低,因为可以从随机突变池中选择稳定的突变。最近,计算方法,如IMPROvER和CompoMug,也已被用于通过建模、生物信息学和神经网络方法帮助预测稳定点突变。利用计算方法显著减少了劳动、成本,并提高了稳定受体构建物的识别速度,然而,选定的稳定突变仍需要通过实验验证。
抗体衍生物或纳米抗体与GPCR的结合可以稳定特定的受体构象,而无需修改天然受体序列,有助于促进高分辨率晶体结构的形成。然而,生产这类抗体工具所需的时间和努力使得这种方法不能被普遍使用。插入融合伙伴可以提供一个稳定的支架,可以增加GPCR的刚性并减少其构象灵活性。此外,融合蛋白增加的表面积可以促进晶体接触的形成,并提高晶体学实验的成功率。然而,与点突变一样,融合蛋白呈现非天然受体序列。在GPCR稳定中使用最受欢迎的融合伙伴是噬菌体T4溶菌酶和热稳定的细胞色素b562RIL(BRIL)。噬菌体T4溶菌酶是一个小而稳定的蛋白质,可以稳定包括GPCRs在内的广泛膜蛋白。另一方面,BRIL是细胞色素b562蛋白的热稳定变体,经过工程改造以提高其溶解度和稳定性。这些融合伙伴可以插入GPCR的不同位置,但最受欢迎的位置是细胞内环3或N末端。
1.6.通过溶解促进GPCRs的膜提取
GPCRs嵌入在细胞膜的脂质双层中;提取它们需要溶解。GPCRs在洗涤剂胶束中的溶解是常见的,然而,这种提取过程有缺点。在溶解过程中,大多数天然脂质从蛋白质中剥离,这可能导致不稳定或非活性的蛋白质产品。
为了克服这一挑战,建议在从膜中提取后采取特定步骤来稳定GPCRs。增加稳定性的一种方法是在前一节中介绍的向GPCR序列本身引入稳定点突变。然而,仔细选择用于GPCR溶解的洗涤剂是另一个重要的因素,可以帮助稳定蛋白质。温和的洗涤剂对GPCR的天然结构影响有限,并且在某些情况下,保留了强烈的天然蛋白质:脂质相互作用。用于结构研究的GPCR溶解最流行的洗涤剂是n-癸基-β-D-麦芽糖苷、n-月桂基-β-D-麦芽糖苷、n-壬基-β-D-葡萄糖苷和月桂基麦芽糖新戊基糖醇。此外,GPCRs通常也在胆固醇半琥珀酸存在下溶解,胆固醇半琥珀酸是一种类似胆固醇的分子,已知对洗涤剂溶解的GPCRs具有稳定作用。
虽然洗涤剂溶解法仍然是药物发现中生产GPCRs的流行方法,但新兴技术提供了一种无需洗涤剂的替代方法,用于从膜中提取蛋白质。其中一种技术是使用苯乙烯马来酸(SMA)共聚物,它们可以自组装在膜双层中,并充当“饼干切割器”,从膜中切除被称为SMA脂质颗粒(SMALPs)的蛋白质/脂质盘(图4)。使用SMALPs允许在天然环境中提取GPCRs,保留它们的结构、功能,并在不需要重新组装到人工脂质环境中的情况下进入下游检测。
根据下游检测的需求,GPCR可能需要以不同的可溶状态呈现,以确保其稳定性或与特定技术(如冷冻电镜(冷冻电子显微镜))的兼容性。所需的GPCR形式可能涉及将GPCR从其溶解洗涤剂转移到两亲聚合物或纳米盘。两亲聚合物(如A8-35或PMAL-C8)像SMALPs一样围绕GPCR形成稳定的脂质带。同样,纳米盘由自组装的膜支架蛋白(由Apolipoprotein A-1工程化)组成,它们环绕GPCR形成一个脂质盘(图4)。然而,GPCR必须在洗涤剂中溶解后重新组装到这些系统中,因为GPCR不能直接被提取到这些系统中。与SMALPs相比,这个较长的过程存在轻微的缺点,因为必须人工添加脂质,而不是利用细胞膜中的天然脂质。此外,在纯化过程中,通常需要将溶解的GPCR从一种洗涤剂胶束转移到另一种,因为最适合溶解和纯化的洗涤剂可能不是最适合下游应用或长期稳定性的。Digitonin和glyco-diosgenin是冷冻电镜分析常用的洗涤剂。
图4. GPCR在膜提取后的不同可溶性呈现形式的例子。左侧的描述说明了每个系统之间的差异,而右侧的详细内容展示了在每种情况下形成活性可溶化剂的化学实体。
荧光尺寸排除色谱法(FSEC)是一种强大的技术,可以作为筛选策略,用于在进行全面纯化之前快速评估GPCR的质量和属性。这种方法提供了一种快速有效的分析GPCR的方法,可以在不首先优化纯化过程的情况下,从粗提物中溶解。FSEC可以快速分析影响GPCR质量和稳定性的各种参数,包括不同的构建设计、用于溶解的洗涤剂、缓冲成分和配体添加。该技术涉及用荧光标签标记感兴趣的蛋白质,该标签可以在尺寸排除色谱期间检测。在FSEC分析期间,蛋白质样品被注入到尺寸排除柱上,该柱根据其分子量分离蛋白质。然后通过荧光检测监测洗脱剖面,并使用所得数据评估GPCR的质量。例如,如果蛋白质以单分散峰洗脱,这表明蛋白质是均匀的并且正确折叠的。另一方面,如果蛋白质以宽峰或多个峰洗脱,则表明蛋白质聚集或错误折叠。
1.7.GPCR生物物理检测
任何药物分子的生物物理评估对药物发现过程都很重要。它能够详细表征所讨论分子的结合亲和力和/或动力学,并以明确定义的方式确认目标参与。这些关键因素使体外生物物理技术成为目标参与的黄金标准,尽管细胞内目标参与技术和技术的快速扩展。传统上,膜蛋白,特别是GPCRs,由于其天然、未修改的构建通常在从细胞膜环境中分离出来时稳定性和活性较差,因此研究起来特别具有挑战性。然而,这种情况在过去十年中随着生物物理技术和GPCR蛋白的制备(见前一节)的进步而迅速改变,并有多个目标使用一系列技术进行研究。与任何目标一样,小分子治疗剂的性质可能使它们更具挑战性,无论是由于低分子量(如SPR)缺乏信号还是可能难以完全表征的初始命中的较弱亲和力。幸运的是,生物物理学家的工具箱从未如此多样化,有许多高度敏感且经过充分验证的技术是该领域的主要技术(如SPR、NMR、LC-MS和ITC(等温量热法))以及提供比既定技术更多优势的更近期的技术,通常与蛋白质的环境有关(例如,细胞内、溶液内/无基质、粗样混合物等)。这些技术都与结构表征(见下一节)紧密相连,这对于片段基础的药物发现项目同样重要,以实现更高吞吐量的结合评估,可以与更低吞吐量但更详细的结构评估相结合。
对于许多用于评估结合亲和力的生物物理技术来说,灵敏度和吞吐量是最重要的进展。例如,现在在合理的时间框架内对数千甚至数万种分子进行生物物理筛选越来越现实,特别是通过SPR,在某些情况下,每周可以完全表征数百种化合物的亲和力和动力学。对于许多筛选管道,吞吐量使项目能够按预期时间表交付,并且能够在多次大型筛选活动(例如,HTS、DEL等)之后生成高质量的生物物理数据是非常宝贵的。低分子量分析物也不再像过去那样具有挑战性,能够使用高通量SPR系统对大型蛋白质构建物(>200 kDa)进行小分子片段筛选。这同样适用于GPCRs,它们通常因表面活性低而受到阻碍,但由于这些进步而变得更加可行。SPR仍然是药物发现管道中的坚定最爱,因为它的吞吐量、灵敏度以及在标准实验中常规提供的信息量,能够表征从mM到sub-pM范围内的分子结合,除了亲和力和目标参与数据外,还可以评估化学计量和其他结合行为(不寻常的动力学、非特异性结合等),这在确认其他技术(如HTS屏幕中发现的)的命中物质时非常有用。话虽如此,生物物理技术的选择应根据所观察的系统量身定制,并且是定义筛选级联的一个组成部分。NMR在评估动态系统或寻找溶液格式中的额外结构洞察方面具有几个优势,并且已在GPCR配体结合研究中使用。像MST(微量热泳动)和DSF(差示扫描荧光测定)这样的技术也提供了无基质的溶液内方法,并且能够应对复杂溶液和/或大分子量目标或难以用生物物理学表征的目标。相关的生物物理平台,如基于细胞的SPRM(表面等离子共振显微镜),是一种正在发展的技术,可以测量小配体和生物制剂在细胞表面原位与GPCRs的结合。SPRM目前不是研究GPCR配体结合动力学的主流技术,尽管已经发表了各种早期采用者的案例研究。
尽管现有的生物物理工具箱很强大,但生物物理检测设计中最重要的步骤无疑是构建本身的设计和准备(见前一节)。许多严重问题通过调整构建长度、突变、溶解条件和标签等因素来解决,甚至在进行生物物理检测之前,可以在检测系统中进行微调。这种针对GPCRs等目标的构建设计和检测设置的协同作用确实决定了并促进了这些目标在生物物理检测中的可行性。
1.8.受体结合检测
配体结合检测是用于询问GPCRs的最悠久的检测格式之一,自20世纪70年代以来就被行业使用。结合检测相对简单,只需要受体来源和合适的配体,通常带有放射性同位素或荧光团的标记。除此之外,这种检测格式可以修改成许多格式,为许多目的提供了丰富的药理学数据。在药物发现环境中,稳健的结合检测可以作为主筛选,用于检测目标GPCR的竞争性和非竞争性配体。这些传统上是在基于滤膜的格式中使用细胞膜制备运行的,但是接近基于检测格式的出现,如闪烁接近颗粒和基于BRET(生物发光共振能量转移)的系统,已经增加了这些检测的吞吐量并减少了手动处理要求。
除了筛选之外,一个经过充分验证的结合检测可以定义标记追踪器或竞争配体的亲和力、结合动力学和结合模式。它们也可以用于询问目标受体的细胞表面表达水平和受体内化。在提出筛选目的的结合检测时需要考虑几个因素:
标记配体和相关试剂可能成本较高,特别是在高通量筛选(HTS)中筛选大量化合物时。
确定合适的标记配体有几个标准,即对目标受体/s具有良好的亲和力和特异性,以及合成路线或标签容忍性的实用性考虑。
对预期目标的配体的了解很重要。结合检测需要一个标记的配体,因此需要一个已知的配体来标记,所以孤儿受体或配体具有较差特异性或亲和力的受体可能使结合检测无法考虑。
这些检测传统上使用放射性标记的追踪配体。氢-3(^3H)是一种具有长半衰期、与固体和液体闪烁剂兼容,并且在化学上与其替代的氢原子相同的同位素,非常推荐使用。碘-125(^125I)是另一种常用同位素,由于其发展成熟且简单的化学性质,在日常肽标记中得到应用。我们发现,使用专业的肽生产和放射性标记组织对肽进行氢-3标记,也能产生一个标记良好的肽,其明显优势是具有12年的半衰期而不是60天,消除了不断重新合成/购买快速衰变碘化配体的需要。最后,使用放射性配体的GPCR结合检测在开发针对GPCR的治疗药物中继续发挥积极作用,最近在2022年批准用于治疗II型糖尿病的GIP受体激动剂Tirzepatide的专利申请中有所描述。此外,其他放射性配体,如PET(正电子发射断层扫描)标记的GPCR配体,在体内受体成像和CNS中的占有率中发挥翻译作用,例如用于多巴胺和5-羟色胺受体药物(见后面的生物标志物部分)。多年来,为了减少对放射性化学药品的处理风险和实际限制,已经开发了几种技术来替代放射性标记配体。这些技术包括荧光强度、荧光偏振、HTRF(如TagLite)、BRET、NanoBRET、流式细胞术和分割荧光素酶系统。荧光GPCR配体可以提供相对受体表达、受体内化、配体亲和力和结合动力学的实时、同时读数,同时保持与放射性标记配体通常相关的稳健信号大小。基于BRET的结合检测近年来变得流行,随着荧光配体和带有荧光标记配体的互补标记或标记受体的商业供应商越来越多。结合检测在行业和我们客户群体的许多药物发现和开发活动中继续发挥关键作用。非放射性系统越来越多地用于体外筛选,而放射性结合检测仍然需求量大,特别是用于证明其在原代细胞和组织中的可转化性。放射性配体有助于确定目标参与、位置、占有率和体内亲和力等属性,其中内源性受体以未修改的形式原位表达。
1.9.基于细胞的检测
为检测类型和筛选级联中的层次选择合适的细胞系统对成功至关重要。主要使用表达重组GPCR的非疾病相关细胞系,如CHO-K1、U2OS、Hela和HEK293,因为它们易于操作、生长特性和广泛可用性。这些细胞系统为使用第二信使检测进行命中识别提供了经过充分验证和稳健的平台。然而,需要注意的是,每种细胞类型内源性的信号和调节蛋白可能会影响检测结果,并导致“系统或细胞类型偏差”,影响测试配体的药理学特性。适当的重组或内源性GPCR细胞模型对于进入体内机制受体-配体和转化研究很重要。另一个重要考虑是是否测量GPCR激活后的近端或远端信号。近端读数将减少由于较少的潜在非靶标介入而产生的不需要的假阳性频率;然而,检测窗口可能小于远端信号。远端信号从受体后信号放大中受益,提供更高的信噪比,然而,假阳性的可能性增加。总体而言,使用精心选择的一系列检测来识别命中,然后在正交检测中进行确认,增加了化合物进展的机会。
1.10.第二信使检测
用于药物发现项目的GPCR基于细胞的筛选检测的经典方法通常测量第二信使的活性,如Ca2+释放、pERK、报告基因表达、1,4,5-肌醇三磷酸(IP3)或IP1积累和cAMP。其他方法包括检测β-arrestin对GPCR的招募或β-arrestin诱导的GPCR内化。通常用于Gq偶联GPCR的第二信使检测测量IP1或Ca2+ i瞬变。广泛使用的IP1检测是一种均质时间分辨荧光检测,它稳健且适用于HTS,虽然试剂成本较高,但检测可以微型化。越来越多地,冷冻细胞可以分配到检测就绪的板上,以避免与连续细胞培养相关的检测运行变化,实现更耐用和可重复的检测。相比之下,Ca2+ i瞬变检测由于其高通量特性和与实时荧光板阅读器(如FLIPR或FDSS)兼容的细胞通透性Ca2+敏感荧光染料的广泛可用性,成为GPCR筛选检测中使用最广泛的技术之一。任何Gi偶联GPCR也与钙结合,从而增加了其作为筛选检测的实用性。除了GPCR靶标外,这些平台还与离子通道和转运体等其他靶标类别兼容,使它们成为药物发现工具箱中有价值的补充。Ca2+ i通量检测的一个重要限制是反应的快速和短暂性质,这限制了对药物MOA的洞察,因为许多缓慢解离的化合物将由于反应的非平衡性质而被视为不可逆拮抗剂。这些方法也可以通过表达嵌合G蛋白(Gαqi5或Gαqo5)或杂种G蛋白(Gα15/16)的重组系统应用于非Gq偶联GPCR,以实现IP1和Ca2+读数,而无需了解G蛋白偶联条件。然而,这带来了风险,即药理活性可能无法从这些重组系统转化为疾病相关细胞系统中的功能检测。GPCR的另一个关键下游读数是测量细胞内cAMP水平。cAMP检测可用于筛选Gs偶联受体(促进cAMP积累)和Gi偶联受体(抑制cAMP产生)。现在有许多技术可用于测量cAMP积累,包括基于抗体的均质检测,如AlphaScreen和HTRF,在药物发现中已广泛使用。另一种方法是使用包含cAMP结合域与荧光或发光蛋白偶联的基因编码生物传感器,如Promega GloSensor,见Kim等人。cAMP生物传感器的优点之一是它们可以用于实时监测cAMP积累,以量化不仅是cAMP的最大水平,还有动力学。ERK1/2检测测量G蛋白依赖和β-arrestin机制的汇聚信号,使其成为初级筛选检测中的常见选择。从许多选项中选择一个稳健的平台是构建高效筛选级联的关键。在选择检测技术和系统时,必须考虑吞吐量、成本和可靠性等关键因素。通常使用如CHO-K1细胞等系统,因为它们易于操作目标基因表达,并且由于仓鼠来源,不太可能表达与目标具有相似同源性的内源性GPCR。用于检测pERK的HTRF均质检测可实现高通量和可重复性;我们已成功使用这种主检测进行了多个GPCR活动。与pERK相关联的一个挑战是由于CHO-K1细胞对播种条件和密度的压力非常敏感,背景较高。为了开发这些检测,我们发现制作大量的冷冻细胞库以最小化传代变化,以及在检测板播种前进行血清饥饿步骤是重要的。
β-arrestin 在 GPCRs 的激活和通过受体内化引起的脱敏中发挥作用,并且是 G 蛋白信号传导的修饰因子。测量 β-arrestin 的招募可以作为主要筛选检测;然而,了解特定的 GPCR 机制和信号偏差是必需的。有多种技术选择可用,如 NanoBit、Tango、β-半乳糖苷酶酶片段互补和 BRET 传感器。这些技术大多使用目标 GPCR 和 β-arrestin 上的亚单位标签,以至于在受体招募时,两个标签部分通过互补被聚集在一起,在底物存在的情况下产生信号。这项技术提供了高信噪比,有利于良好的筛选,并且可以实时测量。
GPCR 内化是 β-arrestin 招募的功能结果,并且已被用于配置配体。这种方法的一个关键优势是不需要事先了解 GPCR 目标信号通路,因此,它有助于为孤儿受体提供帮助。内化可以通过多种检测格式进行测量,例如,我们发现 HiBiT 系统是筛选的稳健方法。HiBiT 内化检测使用细胞外 N-末端 HiBiT 标签对目标 GPCR 进行标记,无论是使用 CRISPR 敲入还是瞬时表达。它依赖于与非细胞通透的 LgBit 在底物存在下的互补以产生发光信号。我们发现内化和 β-arrestin 招募是识别假阳性和建立对主要命中的信心的有用辅助检测。这些检测依赖于低水平的表达,由于技术的灵敏度,建议对瞬时表达进行滴定,以确保目标处于接近内源水平。内化和 β-arrestin 招募都可用于动力学测量,提供对配体激活更深入的机制洞察,但与 HTRF pERK 检测相比,由于信号的持久性,吞吐量更有限。
1.11.G 蛋白生物传感器
GPCR 信号传导主要是通过激活 G 蛋白进行的,其中 GDP 被 GTP 替换在 Gα 亚基的 Gαβγ 三聚体上。这导致 Gα-GTP 和 Gβγ 的活性信号物种。已经有广泛的工具开发来监测这些 G 蛋白的活性,作为激活的直接读数。与可能因途径串扰和信号放大而受到影响的第二信使检测相比,G 蛋白传感器使药理学配置文件基于不同 G 蛋白亚型的结构变化。这些可用于研究与大多数 G 蛋白家族的动态相互作用,包括 Gαi/o、Gαs、Gα12/13 和 Gαq/11。这些检测使用基于 BRET 的传感器,通过 NanoBit 或 BRET 测量异三聚体重排,通过单分子 BRET 测量 G 蛋白-GDP/GTP 交换,通过 MiniG 蛋白 BRET 传感器或 Gβγ/GRK BRET 传感器测量 GPCR/G 蛋白相互作用。
单分子生物传感器使用膜锚定序列融合 Nluc 作为供体,与 YFP 融合肽作为细胞内受体,通过 ER/K α 螺旋连接子连接,以测量细胞内内源性 G 蛋白的激活。这种单分子生物传感器可以在所选择的细胞中表达,具有膜锚定和活性 G-蛋白检测模块(例如,KB-1753)在生物传感器的两端融合。ER/K 连接子有两种构象“弯曲”与“开放”,并且这个比例可以通过连接到连接子两端的蛋白域增加。在非活性状态下,连接子处于“开放”位置,检测到的 BRET 信号很低。然而,当检测模块与膜上的活性 G-蛋白物种结合时,会倾向于“弯曲”构象,将 Nluc 和 YFP 聚集在一起,产生增加的 BRET 信号。这种方法可以应用于原代细胞,如神经元,以了解配体诱导的内源性 G-蛋白调节。整合仅依赖于内源性 G 蛋白表达的传感器消除了使用容易转染的系统(如 HEK293 或 CHO-K1),这些系统通常需要过度表达目标 GPCR。因此,虽然过度表达系统是用于命中发现的标准工具,并且具有显著优势,但在更疾病相关的模型中,通过开发单分子 G-蛋白传感器可能更容易获得高通量替代品。其他团体已经开发了一种传感器,利用原生受体和 G-蛋白揭示配体导向的信号偏差。虽然这些技术在发现配体偏差和近端活性测量方面具有巨大潜力,但目前尚不清楚这些技术在 HTS 命中发现活动中的扩展程度如何。常见 GPCR 检测和技术的总结表可以在补充 1 中找到。
1.12.GPCR 目标参与
GPCR 参与的一个潜在方法,无论是用于命中发现还是领先优化,是 HiBiT 或其他分裂报告标签的适应,如 Pro-link 用于细胞热漂移测定(CETSA)。这些分裂报告克服了对可靠检测抗体的需求,这是 GPCR 目标的常见问题,并且已用于 HTS 活动。虽然原生 CETSA 对 GPCR 来说比其他类别的目标更具挑战性,但有使用原生抗体进行一些 GPCR 目标的先例。原生 CETSA 方法对跨膜蛋白的挑战很明显,并且不太可能广泛适用于所有 GPCR。然而,使用替代检测系统而不是依赖于基于抗体的检测可能有助于确定这些挑战是由于缺乏合适的抗体还是 GPCR 目标的属性。当存在重组蛋白生产的固有挑战、没有可靠的抗体和缺乏 Nano-BRET 方法的探针时,分裂报告检测系统可能提供了一种前进的方式。我们发现使用易于转染的 HEK293 细胞以瞬时方法是最短时间内成功检测开发的最短途径。检测灵敏度直接与表达水平相关;因此,表达滴定对于构建稳健的筛选检测非常重要。利用反向转染直接到检测板或用于创建冷冻细胞,已经实现了中等吞吐量筛选检测的执行。
1.13.药物发现中的现象学检测
1.13.1.我们如何测量新药的效力?
如上所述,我们选择选择新化合物的检测类型根据我们试图回答的问题差异很大。检测可以从非常简单的体外结合或信号检测,如前所述,具有高灵敏度和吞吐量的优点,到更生理相关的但吞吐量低的体内疾病模型。
使用原代人类细胞和现象学筛选可以弥合这两个极端之间的差距,提供具有足够吞吐量的检测,允许筛选大量化合物,结合目标在疾病相关细胞系统中的内源性表达。原代人类细胞中的现象学检测将使我们能够模拟复杂疾病的多个方面,因此,与动物模型相比,更有可能预测临床结果。
1.13.2.现象学筛选的重要性
现象学检测测量细胞在潜在药物处理后的行为变化。一些常用的示例包括细胞移动或增殖、介质释放和细胞分化。这些复杂的检测最好在疾病相关的细胞系统中监测,例如原代人类细胞,目标在(病理)生理水平上表达,并与正确的辅助蛋白共定位。表达水平和细胞环境对 GPCRs 尤为重要,因为不同类型的细胞以不同水平表达不同的 G 蛋白亚型,这可能会改变 GPCRs 在简单过度表达的细胞系系统中的信号传导。
GPCRs 的一个关键特征是受体激动剂表现出效力和效力,这与受体表达、储备和信号放大内在相关。例如,高效力或完全激动剂在低受体占据下产生最大效应,无论所研究系统的受体表达水平如何。相比之下,低效力或部分激动剂必须占据高达 100% 的可用受体才能启动响应。在低表达系统中,即使占据所有可用受体,部分激动剂也只能产生亚最大响应。随着受体表达的增加,部分激动剂可以占据的受体更多;因此,它们产生的响应增加,直到达到系统最大值。这一发现在许多 GPCRs 的领域已经得到证明,例如,β2 肾上腺素能受体激动剂沙丁胺醇和特布他林在支气管平滑肌中是完全激动剂,那里有大量的受体储备,但在受体表达低得多的肥大细胞中是部分的。为了获得激动剂效力的准确测量,我们必须仔细选择与我们感兴趣的疾病最相关的细胞系统。
激动剂在探索急性或慢性反应时也可能表现出不同的药理特性,因此,识别最适合的检测方法和时间点以表征药物发现中的新化合物至关重要。简单的第二信使检测并不总是能预测在更慢性的表型读数中的内在活性。例如,在人类原代肺成纤维细胞中已经证明,一系列Gs偶联GPCRs产生的cAMP的量并不能预测相同配体抑制成纤维细胞增殖或分化的能力。当选择最理想的药物特性以针对急性生理反应(如哮喘中的支气管扩张)与慢性反应(例如,纤维化中的气道重塑)时,这种细胞类型差异是一个重要的考虑因素。
1.14.药物发现中的细胞成像
根据您希望测量的细胞反应,有各种方法可以测量表型变化。这些可以从简单的检测试剂盒,量化分泌到细胞外介质中的蛋白质,到原代细胞中更复杂的多目标标记。免疫荧光显微镜是用于探索细胞表型反应的更强大的工具之一。这种方法允许我们监测细胞期望的物理特性的变化,并为我们提供了可以指示我们的化合物可能介导的细胞健康意外变化的细胞的视觉表示。
传统的显微镜是手动的、通量低、耗时且技术密集的,使其不适合药物发现筛选。高内容筛选或高内容成像(HCI)结合了强大的显微镜和基于板的检测,允许我们执行自动化和定量的荧光显微镜检测。HCI旨在通过多波长成像和自动化图像获取,尽可能多地获取有关细胞、球状体或类器官的功能和形态变化的数据。它允许在高通量检测中单细胞或多细胞成像,并且适用于微型化和自动化。因此,它已成为GPCR药物发现工具箱中有价值的工具。
HCI旨在通过多波长成像和自动化图像获取最大化数据捕获,这意味着我们可以从单独的载玻片转向96/384孔板。由于捕获的数据量很大,它还利用了自动化图像处理,其中多参数算法可以使用机器学习识别一系列形态特征。这种自动化图像获取与分析的整合允许产生高度复杂的定量数据。有许多HCI平台可用,如Molecular Devices ImageXpress、Perkin Elmer Opera Pheonix、Yokogawa CV8000或ZEISS CellDiscoverer。每个系统将为光源、放大倍数以及执行宽场或共焦成像的能力提供不同的选项。
HCI可以利用任何发出荧光信号的标签或探针。只要仔细选择探针,使激发/发射光谱不重叠(图5),不同的探针就可以在相同的视野内多重化。甚至最简单的检测通常也涉及某种形式的多重化。通常会包括像Hoechst这样的核染色,以便准确识别细胞、分离和检测量化。
图5. 可用于使用高内涵成像(HCI)进行多重检测的不同荧光探针的例子。
图表展示了通过选择不重叠的激发和发射光谱多重化不同荧光团的能力。模拟的光谱(Ex/Em波长)显示为A)350/450 - 蓝色,B)530/560 - 绿色和C)647/700 - 远红色。
任何细胞蛋白都可以使用HCI进行监测,只要有足够的选择性初级抗体可用。这些可以与荧光标记的二抗体结合使用,以高度敏感的方式准确标记细胞蛋白。这种方法允许监测目标蛋白的表达和定位,以及一些翻译后修饰,如蛋白质磷酸化。如果灵敏度不是问题,初级抗体可以被荧光染料标记,许多这样的标记试剂盒可以商业获取。在没有选择性抗体的情况下,像许多新型GPCRs一样,我们可以寻找替代方法来标记蛋白质,如荧光配体或直接将荧光基团添加到受体或目标蛋白上。这种方法可以使用配体指导GPCRs与荧光标签在活细胞中的共价标记,允许我们监测内源性受体而不是依赖标记受体的过度表达。这些方法可以监测受体与配体的结合以及这些受体的细胞位置。
除了抗体标记,我们还可以利用荧光染料有选择性地标记细胞内的不同结构和细胞器。有可用于标记质膜、细胞质、溶酶体和线粒体的例子,仅举几例。这些中的许多都有不同的颜色以允许多重化,并且有pH敏感染料,一旦它们到达细胞内囊泡的酸性环境,如晚期内体和溶酶体,就只会发出荧光。例如,法尔定常用于选择性染色F-肌动蛋白,这允许监测肌动蛋白骨架的重排,这是形状变化、伤口愈合、细胞运动和细胞粘附的典型特征。
HCI还可以利用基于FRET的生物传感器探索细胞内信号事件(图6)。最近,这些已经被靶向到不同的细胞区域,以允许详细成像亚细胞隔室。因此,这些类型的生物传感器将使我们能够探索GPCRs下游信号的动力学和隔室化,有助于解释可能发生在仅有几个G蛋白下游的不同表型响应的复杂性。
图6. 一系列高内涵成像(HCI)的例子,展示了 (a) GPCR驱动的成纤维细胞迁移 ,(b) 细胞内ERK磷酸化 和 (c) 细胞染色 。
结合这些标记技术的一些或全部,允许我们捕获细胞的特定表型或形态特征的详细图片,这个过程被称为细胞绘画。高通量成像细胞被多达6种荧光染料染色,允许我们尽可能多地“标记”细胞的不同区域(见图6)。自动化图像分析然后使用这些图像识别数百到数千个不同的细胞形态特征,提供高度详细的表型特征。
高通量表型和基于成像的检测,特别是在原代人类细胞中,可以帮助我们以更生理和病理生理的环境了解我们的GPCR目标和新化合物。这种HCI过程允许我们在漫长的体内研究之前更好地分类化合物,因此有助于减少药物发现中晚期失败。
1.15.作用机制
阐明药物的MOA涉及将实验数据与受体药理学机制模型进行比较。简单的MOA研究从更具挑战性的评估弱、命中化合物开始,到更复杂的分析,具有更丰富、高效的领先化合物。由于药物可能通过与同一受体的相互作用在不同组织中产生不同的效果,因此最有用的模型描述了药物的系统独立属性。这些模型使药物发现研究人员能够使用测试系统中的实验数据来预测化合物在目标组织中的活性。
在药物发现计划中识别的GPCR配体可能作为激动剂、中性拮抗剂、反向激动剂或别构调节剂。激动剂是激活受体以产生生理反应的配体,而中性拮抗剂结合到受体上并不启动反应,从而阻止激动剂的作用。反向激动作用可以发生在具有组成活性的系统中 - 受体的自发活性 - 这可以通过结合反向激动剂来逆转。组成活性和反向激动作用的现象可以使用双态模型来解释,在该模型中,受体可以存在于活跃或非活跃的构象中。根据这个模型,偏爱活跃受体状态的配体将促进激活,并被表征为激动剂。相反,反向激动剂对非活跃状态的亲和力大于活跃状态,并破坏自发受体激活。与活跃和非活跃状态具有相等亲和力的配体将表现为中性拮抗剂。
1.15.1.激动剂
定义细胞系统中激动剂浓度-反应曲线的参数取决于系统独立因素和系统依赖因素。因此,应在具有不同敏感性的多种细胞类型中表征新化学物质。在命中识别筛选中检测激动剂的理想场景是高度敏感的,以揭示对弱激动剂的反应。相反,为了在命中到领先或领先优化计划中区分激动剂,应该测量不太敏感的细胞反应,以便化合物可以被表征为完全或部分激动剂。
根据细胞系统中的最大反应,激动剂可以被描述为完全或部分。完全激动剂产生等于系统最大值的最大反应,而部分激动剂即使在高浓度下也只能产生亚最大反应。已经描述了一个配体子集作为超激动剂,相对于内源性配体。超激动剂能够在相同检测中产生大于内源性激动剂的最大反应。两个具有相同机制的完全激动剂的EC50值的比率在所有它们产生完全激动作用的细胞系统中应该是恒定的。因此,通过计算等效活性效力比,可以简单地量化完全激动剂的活性。可以从浓度-反应曲线中获得更多关于部分激动剂的信息,因为相对效力较低的激动剂表现出EC50值接近激动剂-受体复合物的平衡解离常数(KA)。
因此,部分激动剂可以根据效力和最大反应进行排名,分别表示亲和力和效力的等级顺序。将激动剂与反应联系起来的定量关系由Black-Leff操作模型描述。
在最大反应Em的系统中,对激动剂浓度[A]的反应由平衡解离常数KA和系统中激动剂的效力决定。后者由换能比τ给出,定义为R0/KE,其中R0是总受体浓度,KE是产生半最大效应的激动剂-受体复合物的浓度。简单来说,τ是结合亲和力和功能性效力(效力)之间的差异。因此,系统中激动剂的效力取决于激动剂的内在效力和细胞系统的敏感性。
从Black-Leff操作模型拟合中导出的系数τ/KA,衡量激动剂诱导特定反应的能力。对于完全激动剂来说,由于可以无限组合τ和KA值来拟合数据,因此很难从浓度-反应曲线中确定效力和亲和力。然而,可以确定τ/KA比率的唯一值。当Hill斜率(n)为1时,系数τ/KA等同于Emax/EC50相对活性,其中Emax是激动剂的最大反应,作为系统最大值的分数。通过计算ΔLog(τ/KA)或ΔLog(max/EC50)(当n≈1时),可以对激动剂进行排名,给出一个系统独立的指数。
GPCR配体能激活多个途径,具有不同的效力和效力,这种现象称为偏向信号。这是配体依赖性地激活特定途径而不是其他途径,例如,激活多个G蛋白依赖性信号途径,或G蛋白依赖性信号对arrestin介导的信号修饰。如果它们由不同的信号途径介导,偏向信号可能在治疗上被利用,以实现药物效力,同时避免不良反应。准确量化偏差在技术上可能具有挑战性,因为许多检测伪影可能会使偏向信号的解释复杂化。这些包括检测设计和被测量反应的动力学差异,途径之间的信号放大或受体储备的差异,以及意外的非靶标效应。然而,通过仔细控制这些因素,我们可以使用不同的模型为配体分配偏差因子,以了解多个途径之间的信号关系。获得真正的偏向药物以及理解信号偏差的治疗潜力的困难由所谓的G蛋白偏向μ阿片受体激动剂Oliceridine所说明。例如,Oliceridine在临床试验中是一种可证明的有效的镇痛药,但最初观察到的治疗窗口和显著偏差可能可以解释为在具有高受体储备的系统中部分激动剂放大反应。
这些模型已在最近的一套指南中描述,其中配体偏差可以通过比较不同途径中测试配体与参考配体的Log(τ/KA)或相对活性值来量化。然后由偏差因子10ΔΔlog(τ/KA)或10ΔΔlog(max/EC50)表示配体依赖性的途径激活差异,这是一种无论来自配体亲和力还是效力差异的偏差度量。
1.15.2.拮抗剂
分子可以通过两种机制拮抗激动剂的反应。正性拮抗作用发生在拮抗剂与正性激动剂结合在同一位点并阻止激动剂与受体结合时。相反,别构拮抗剂在特定位点结合并通过蛋白质的构象变化与激动剂相互作用。
拮抗作用的性质可以进一步用行为来描述,即可克服或不可克服,竞争性或非竞争性。这种行为描述取决于观察到拮抗作用的检测条件。在竞争性正性拮抗作用的情况下,非常高浓度的激动剂仍然能够产生最大反应;即,拮抗作用是可以克服的。在这种情况下,可以执行Schild分析以确定结合亲和力。在此检测中,进行浓度-反应曲线,随着拮抗剂浓度的增加,导致激动剂曲线平行右移。通过Schild回归得到的斜率因子的一致性来确定竞争性拮抗作用模型的符合性。检查激动剂-拮抗剂相互作用的最严格方法是使用修改的Gaddum-Schild方程进行全局非线性回归。
当我们为药物发现对新化合物的MOA进行分类时,对所有化合物执行完整的Schild回归通常不是时间高效或实际的;因此,我们可以依靠使用竞争结合检测来确定IC50和Ki。对于这种分析,我们只需要在控制追踪配体存在的情况下对每个未知化合物执行一次竞争检测,以推导IC50值。IC50是一个依赖于使用的追踪配体的浓度和亲和力的经验值;因此,我们使用Cheng-Prusoff方程将这个值转换为Ki。通过这种方式,我们可以准确地测量许多新化合物的结合亲和力。
相反,非竞争性拮抗行为是不可克服的。这种行为可能源于伪不可逆拮抗剂占据正性位点或别构效应。在高浓度的拮抗剂存在下,激动剂无法产生其最大反应,导致拮抗剂浓度依赖性地抑制激动剂浓度-反应曲线。
1.15.3.别构调节剂
别构调节剂通过结合到与正性结合位点空间上不同的位点来影响GPCRs。因此,它们可能为药物发现提供优势,通过赋予增加的受体和受体亚型选择性。
别构调节的一个独特特点是效果的饱和性,即,一旦所有别构位点都被占据,即使在更大的调节剂浓度下也不会观察到对亲和力或效力的进一步调节。对于别构拮抗剂来说,这可以被视为一个曲线Schild图,其中连续的激动剂浓度-反应曲线在高调节剂浓度处聚集在一起。别构配体可能调节正性配体的亲和力和/或效力,也可能具有直接激动作用。因此,在测量受体功能的检测中可能存在许多不同的别构效应。所有这些效应都由功能性别构模型描述。在一系列调节剂浓度存在下拟合完全激动剂浓度-反应曲线,可得到调节剂亲和力(KB)以及亲和力(α)和效力(β)的协同因子的系统独立参数值。
在药物发现中,正性别构调节剂(PAMs)的一个优势是它们区分激活和非激活受体状态,从而增强对正性激动剂的反应。通过这种方式,PAMs可以通过在适当的时间和适当的地点增强现有反应来微调对内源性配体的反应,因此可能比激动剂具有更广泛治疗窗口。PAMs可以使用传统的激动剂筛选方法检测;然而,新化合物在低浓度存在下进行测试,例如,正性激动剂的EC30,以确定它们是否可以增强其反应。
别构和正性拮抗剂可能在功能性基于细胞的和生化结合检测中共享一些行为。根据实验条件,如果在使用选择性探针来表征测试化合物时没有观察到探针依赖性,或者在测试浓度的溶解度限制范围内没有达到饱和性,NAM可能被错误地解释为正性拮抗剂。可以通过竞争关联检测或放射性配体解离动力学检测来研究疑似别构调节剂的机制。观察到的激动剂-受体复合物的解离速率常数的变化可以归因于别构效应。
有许多不同的机制,潜在药物可以通过这些机制调节GPCRs的活性,理解治疗特定疾病最合适的机制可以帮助我们制造更好的药物。早期选择所需的机制可以帮助设计相关的初始筛选检测,如PAMs,或选择适当的机制研究,如偏向配体。新化合物的不同药理属性的量化可以进入DMTA循环,允许我们通过领先优化阶段推进更好的化合物。除了标准机制研究外,我们还可以利用细胞绘画等方法获得关于化合物MoAs的非假设驱动信息。如前所述,可以开发详细的细胞配置文件,使用HCI然后用于检测不同实验化合物处理的单个细胞中的微妙差异。这个过程可以识别靶标,测量毒性,将化合物分组到功能途径中,识别作用机制或信号途径中的微妙差异,或识别疾病特征。通过设置使用非假设驱动过程的研究,我们可以最大化新发现的机会。总之,将详细的MOA研究纳入药物发现过程可以帮助设计和优化化合物,以开发更有效的治疗方法。
1.16.GPCR结构生物学
GPCR的成功结构引导药物发现在很大程度上可以通过常规提供实验结构来辅助,这些结构的分辨率允许对结合的小分子进行明确建模。然而,GPCR的结构确定通常是一个漫长而昂贵的过程,并且比可溶性蛋白靶标的失败率更高。因此,在一项运动的开始时,必须考虑GPCR结构的成本/收益分析和可交付的可行时间框架。我们认为,投资于目标GPCR的结构是值得的,因为如果成功,它有助于药物化学优化化合物并提供有关受体结构生物学的宝贵见解。我们认为,这项投资的时机最好是在HTS等命中发现活动之前或同时进行,以便有益于指导SAR。
尽管GPCR的结构生物学具有挑战性,但GPCR结构存入蛋白质数据银行(PDB)的速度正在加快,并且所达到的分辨率一直在提高(图7),这意味着这项任务变得越来越可行和常规。有几个因素促成了GPCR结构生物学的成功,尽管它们是具有挑战性的目标。这些因素包括在构建设计、表达系统以及用于在样本准备过程中溶解和稳定受体的洗涤剂方面的技术进步,正如前面所涵盖的。此外,结构解决方案技术本身也有许多关键技术进步,这些技术在获得GPCR结构方面也起到了重要作用。
在左侧,散点图显示了随时间存入PDB的GPCR结构的累积数量。在右侧,散点图显示了随时间存入PDB的GPCR结构的最大和平均分辨率。
广义上,有两种技术可以确定GPCR的结构:X射线晶体学和冷冻电镜(Cryo-EM)。大部分情况下,由前者解决的是拮抗剂结合/非活动状态结构,而与异源三聚体G蛋白复合物的激动剂结合/活性状态结构则由后者解决。结构技术的的选择对构建设计、表达和纯化过程有很大的影响,因此有助于在项目开始时就知道要针对的理想模式是什么。在2018年之前,GPCR结构主要是通过X射线晶体学解决的。传统的蒸汽扩散晶体化方法已用于解决34%的GPCR晶体结构。另一方面,由Caffrey实验室开创并专门设计用于膜蛋白晶体化的脂质立方相(LCP)方法已用于解决65%的GPCR晶体结构。LCP方法协助晶体化的能力归因于其能够通过在疏水和亲水环境的双相混合物中容纳膜蛋白,提供适合晶体成核和生长的环境。然而,由于蛋白质晶体化需要蛋白质样品处于稳定和刚性状态,迄今为止通过X射线晶体学解决的95%的GPCR结构是A家族GPCRs,它们比其他家族成员更容易晶体化。
通常,在第三代同步辐射X射线源的微焦点光束线上收集LCP晶体的衍射数据。这些光束线旨在产生直径在1-10微米范围内的高度聚焦的X射线束,这允许将高X射线束通量传递到非常集中的区域,允许从GPCR常见的非常小和弱衍射晶体收集数据。由于从这些晶体收集衍射数据需要高X射线剂量,必须从许多晶体收集小楔形数据,以避免由于辐射损伤导致数据质量严重下降。然后合并多个数据集以记录一个完整的数据集,从而可以解决GPCR结构。
串行飞秒晶体学,自2013年首次产出GPCR晶体结构以来,使用X射线自由电子激光器(XFEL)而不是同步辐射源进行晶体学数据收集。这种方法已用于解决迄今为止许多公开可用的GPCR晶体结构。串行飞秒晶体学可以被视为典型X射线晶体学工作流程的扩展,因为数据收集之前的步骤没有不同,直到晶体化成功后。XFELs产生超高强度、超短X射线脉冲,这些脉冲非常亮和相干,它们的使用在研究GPCR结构方面比其他X射线源有几个优点。首先,超短X射线脉冲可以用来捕获不同方向上多个单晶的衍射图案,在X射线诱导的辐射损伤之前捕获衍射图案。第二,XFELs产生极高强度的X射线,这使得能够从以前太弱而无法产生有意义数据的小晶体收集衍射数据。最后,用于串行飞秒晶体学的LCP晶体不需要像在同步辐射X射线衍射实验中那样收获和冷冻,因为在串行飞秒实验中,LCP晶体直接在它们生长的LCP中喷射过去。因此,串行飞秒晶体学可以帮助解决来自LCP中极小晶体的GPCR结构,即使晶体化条件的优化无法产生足够大和质量的晶体,以便使用同步辐射数据源收集数据,即使在微焦点光束线上。
冷冻电镜是解决蛋白质分子结构的另一种技术。冷冻电镜自20世纪60年代以来一直在发展,然而,在过去十年中经历了一系列快速发展,将焦点从X射线晶体学转移开,作为蛋白质结构解决的主要方法。这场所谓的“分辨率革命”是由几个重大进步推动的,包括直接电子探测器的开发、自动化数据收集软件、改进的运动校正和CTF校正算法、复杂的粒子选择方法、单粒子分析软件包,以及对电子显微镜基础设施的普遍改善。自2018年以来,冷冻电镜已超过晶体学,成为获取包括GPCRs在内的许多蛋白质类别结构信息的首选技术(图7)。
图7. 提交至蛋白质数据银行(PDB)的GPCR结构的时间线。
在冷冻电镜实验中,不是记录衍射图案,而是将入射电子束通过在一层玻璃态冰中冷冻的蛋白质样品传递,其中产生的散射电子被放大到探测器上,形成图像。从一系列得到的微观图像中,将不同方向上的单个蛋白质粒子的图像平均在一起,首先构建2D模型,然后通过计算过程构建3D模型。冷冻电镜的优势在于不需要目标蛋白晶体化,并且成功结构解决所需的蛋白质量可能比晶体学运动所需的少10-100倍。此外,使用冷冻电镜,与GPCR结构生物学相关的更顽固的挑战,如它们的不稳定性和差表达和纯化产量,对于结构解决的障碍较小。
从冷冻电镜数据重建三维结构涉及组合成千上万到数百万在玻璃态冰中不同方向上观察到的粒子。这个过程依赖于处理软件正确识别或“选择”来自信号-噪声比非常低的图像中的粒子,并准确分配该粒子的方向。因此,有一个更低的尺寸限制,在这个限制下冷冻电镜是不可能的,因为粒子不再能成功选择。出于这个原因,大多数GPCR的冷冻电镜结构是GPCR:G蛋白复合物和越来越多的GPCR:arrestin结构,因为这些复合物比单独的受体呈现更大的粒子。然而,可以成功解决的粒子尺寸下限一直在变小,一些最新的膜蛋白结构成功解决的尺寸只有60 kDa。
在GPCR:G蛋白复合物中,GPCR必须处于活性状态,这意味着它们是激动剂结合状态的结构,而不是GPCR的非活性/拮抗剂结合状态,这在X射线晶体结构中更为常见。除了异源三聚体G蛋白复合物外,通常使用通用纳米抗体或Fab片段,它们结合并稳定G蛋白亚基界面之间的界面。例如,纳米抗体35(Nb35)是一种构象特异性纳米抗体,结合G⍺和Gβ亚基之间的界面(图8)。
图8. GPCR:G蛋白复合体的冷冻电镜结构示例。
1.16.1.腺苷A2a受体(PDB: 6GDG)
在冷冻电镜实验中增加目标蛋白粒子尺寸的替代策略是使用源自抗体的结合伙伴。最近几篇出版物突出了使用Fab抗体片段的效用,这些片段识别GPCR-BRIL融合的BRIL部分,以协助这些蛋白质的结构表征。因此,在涉及GPCR的药物发现运动中,追求X射线晶体学与冷冻电镜作为首选结构解决方法的明显区别已经变得模糊。然而,如果趋势持续下去,即使在表征小分子结合的情况下,也可能会有越来越多的GPCR目标通过冷冻电镜来解决。
1.17.药物化学见解
在本节中,我们将首先探讨前面描述的GPCR结构信息如何帮助指导药物化学优化药物,然后继续讨论一些特定的拮抗剂SAR和常见结构基元,以及别构和偏向配体SAR和化学的例子。
1.17.1.结构引导的药物发现
GPCRs是动态的信号结构,配体结合口袋可以表现出相当的灵活性以适应配体,这使得确定和建模相关受体-配体结构具有挑战性。然而,随着我们获取相关和多个GPCR-配体结构的能力的提高,以及计算药物-受体柔性对接方法的速度和预测性的提高,将配体诱导的受体构象变化纳入药物发现的能力得到了改善。
使用稳定的mGlu5受体,Cristopher等人进行了基于片段的结合检测以鉴定命中。受体-化合物复合物的X射线晶体结构允许合理化SAR,导致选择一个化合物进行进一步开发。此外,鉴定的化合物结合到mGlu5受体的一个别构口袋内。随后,Christopher等人表明不同的mGlu5配体可以通过一系列诱导拟合采取不同的结合构象。除了配体结合模式分析外,还考虑了结合位点中的水网络。这种方法对于开发GPCR别构调节剂非常有利,因为理解通过在不利环境中去除水分子以及保持与有利水分子的相互作用的相互作用对水分子的影响可以提高结合亲和力,这通过合理的SAR可能更难实现(图9)。
图9. HTL14242是通过基于片段的筛选和使用结构基础药物设计(SBDD)优化发现的。化合物16和19被开发出来,通过稳定结合口袋中的水网络来合理解释效力差异。a 来自激动剂诱导的磷脂酰肌醇水解测定的IC50值。
增加在激活和非激活状态下解决的GPCR结构数量将有助于未来的药物发现工作,为计算同源建模提供更多新颖的模板(见图7)。这些受体-配体结构可以帮助进一步发展通过深度学习或神经网络改进结构预测的人工智能(AI)技术。Deepmind开发的AlphaFold在2021年发布,提供了超过2亿个蛋白质结构模型。这项新技术导致了AlphaFold生成的模型与实验结构的比较。通常,由AlphaFold开发的GPCR模型的三级主链特征非常接近;蛋白质侧链的方向显示出最大的差异,侧链RMSD(1.90 ± 0.58 Å)。这可能是由于结合口袋的疏水塌陷导致AlphaFold结构中更小、更窄的结合口袋。
在AlphaFold和AlphaFold 2的基础上,Heo和Feig制定了一个协议,允许预测GPCR的活性和非活性状态。历史上,许多GPCR X射线晶体学数据集中,PDB数据库中非活性状态的数量过多。Heo和Feng建立了一个新的协议,使用AlphaFold 2并引导它到所需的激活状态。Lee等人的工作提供了一些当前的指导方针,介绍如何使用深度学习开发的GPCR结构模型,例如AlphaFold。两个主要组成部分是(1)选择正确的模型,使用正确的激活状态,以及(2)通过受体柔性对接使用适当的对接选项。
图10. 例子表明,简单的配体修饰可以导致不同的GPCR药理学。
1.17.2.配体的简单修饰
小分子在给定GPCR上的功能性活性可能受到小的结构变化的显著影响。Dosa和Amin在2016年对这些变化的深入审查突出了一些最近的例子,其中小的结构变化显著影响了GPCR药理学。在不同的配体修饰中,Dosa和Amin观察到,在71%的情况下,结构变化γ位置到氮,或更近,导致配体功能从激动剂转变为拮抗剂或反之亦然。这些例子包括:(a) 在CCR3拮抗剂的哌啶环的3′位置的甲基化,增加了结合但转变为激动剂。(b) 在一系列α2-肾上腺素受体激动剂中,N-甲基苯胺向二甲胺的甲基化保持了结合亲和力(Ki),但将机制从激动剂转变为拮抗剂。在这种情况下,SAR并不总是在系列中一致,但显示了小的结构变化可以对α2-肾上腺素受体的功能产生显著差异。(c) 将 pergolide 和 Oopergolide 的 N-丙基基团修改为 N-甲基,将功能效应从5-HT2B激动剂转变为拮抗剂。有趣的是,pergolide 和 6-methpergolide 仍然是多巴胺D2受体的激动剂。
1.17.3.特权支架
特权支架是提供特定属性的化学亚结构,并且在药物化学的情况下,这是特定蛋白质类别的结合基元。特权支架通常是半刚性的,可以通过疏水相互作用结合到疏水区域,而不会崩溃配体的三维结构。特权支架自1988年以来就已经存在,当时Evans等人提出,仔细调节特权支架将有助于找到GPCRs的新型激动剂或拮抗剂。在GPCR家族中,有一些众所周知的特权结构,了解这些结构可以帮助将命中转化为领先化合物。在修改特权支架时应谨慎考虑,以及对配体的整体物理化学性质的影响,例如,通过确保在优化过程中保持配体的亲脂性。GPCRs的一些特权支架示例包括联苯、螺哌啶和苯二氮卓。
图11. 阿立哌唑,一种上市的D2受体部分激动剂,以及一些含有吲哚的类似物,它们在信号偏向性上有不同的效应。
1.18.偏向信号
配体偏向的概念自1995年以来就已经存在,基于不同配体可以通过相同的GPCR通过偏向不同途径的受体后信号(例如,G蛋白偶联反应和β-arrestin的调节)影响一系列细胞功能的想法。在超过30个GPCRs中观察到明显的偏向信号,Tan等人对这一领域的综述提供了不同配体结构和信号通路影响的深入分析。一个关于小配体结构变化如何影响信号通路的例子是一系列多巴胺2受体(D2R)激动剂。利用GPCR结构和机制数据,建立了一个假设,即修改Aripiprazole的二氯苯环为吲哚环系统可以导致与跨膜螺旋的不同相互作用,从而影响信号通路。N-H吲哚化合物1在Gi/o检测中具有与β-arrestin 2招募检测相似的效力,计算出的β-arrestin偏向因子为2.5。简单的吲哚氮甲基化导致配体化合物2在Gi/o检测中无效,但在β-arrestin 2招募检测中仍然活跃,尽管效力有所降低。进一步的甲基化α到吲哚氮化合物7提供了一个在Gi/o检测和β-arrestin 2招募检测中都活跃的配体,对G蛋白信号有显著的偏向,由β-arrestin调节。这是第一个使用基于GPCR结构信息的合理设计生成偏向GPCR配体的例子。
1.19.别构调节剂
GPCRs的别构调节剂结合在与天然配体的正性结合位点不同的位点。别构调节剂修改受体的功能效应的能力,无论是在天然配体存在下积极还是消极,可以提供一种探索潜在的差异化效力、受体选择性和与正性配体相比改善安全性的方法。在GPCR X射线晶体学衍生结构中发现了几种不同的别构结合位点,包括GPCR的细胞外侧、跨膜螺旋和脂质双层之间的结合位点,以及TM 6和7细胞内侧区域底部的结合位点。通常在别构药物化学项目中,配体的结构变化相对较小,活性变化剧烈,使得构建结构活性关系变得困难。
GPCRs的新型别构调节剂的发现是一个不断增长的领域,以下是一些最近发表的别构调节剂的例子,包括1H-咪唑[4,5-c]喹啉-4-胺类似物作为A3腺苷受体的PAMs,V-0219,一种1,2,4-噁二唑-哌啶PAM的胰高血糖素样肽-1受体,噻吩[2,3-d]嘧啶类似物作为Mas相关G蛋白偶联受体X1的PAMs,chromane CVN636 mGluR7的别构激动剂,苯乙基脲类似物作为大麻素类型1受体的NAMs,三唑苯酰胺类似物的mGlu7和噻吩[2,3-d]嘧啶类似物作为D2受体的NAMs。
在最近的一项研究中,披露了一种新型的多巴胺D1受体正性异构调节剂(PAM)。传统的正性D1激动剂曾被用于治疗帕金森病、精神分裂症、阿尔茨海默症以及其他形式的痴呆症;然而,这些正性配体存在一系列缺点,包括对其他多巴胺受体的选择性差、药代动力学特性差;“钟形”剂量反应;耐受性和脱敏;包括低血压在内的耐受性问题。通过针对D1R的正性异构调节,希望能够在不产生正性配体相关不良影响的情况下提高内源性多巴胺的效力。令人鼓舞的是,已有两种D1R PAMs进入了针对帕金森病和精神分裂症的临床开发。
D1R的命中是从在稳定表达D1R的神经母细胞瘤细胞系中进行的cAMP积累读数的PAM筛选的小化合物库中鉴定出来的(图12)。对双苯基的取代进行快速评估,导致在cAMP积累检测中测试了一系列化合物,表明在2位具有氟甲氧基团提供了最有利的活性。同样揭示了在3位具有醛基导致效力下降,与4位相比。总体而言,化合物3具有最佳的特性。最初的命中在cAMP检测中没有反应,表明该化合物作为PAM没有固有的激动剂活性。进一步的放射性配体受体结合研究表明了正性异构效应。尽管化合物3具有良好的渗透性和低hERG抑制作用,但血清稳定性差、溶解度差和高蛋白质结合意味着需要一种具有更好体外PK特性的化合物。亚磺胺类似物(化合物26)显示出显著的效力提高,并且对这种化学类型的进一步研究没有显示出进一步的效力提高。进一步的研究证实亚磺胺类似物作为PAM,ADMET分析显示比化合物3具有更优越的特性。由于亚磺胺基团是手性的,合成了两种对映体,R和S对映体在D1R上具有可比的效力。
图 12. 从最初的醛类命中发展出D1受体正向别构调节剂(PAM)。
简要地,为了完整性,一个最近报道的NAM的例子是GLPG1205,由Galapagos开发的GPR84的NAM。GPR84由中链游离脂肪酸激活,通过Gi/o途径信号传递,与炎症性疾病相关。使用cAMP检测进行了200 K化合物的HTS,GTPγS检测用于命中验证。在这个HTS中,鉴定了一个具有亚微摩尔效力和良好物理化学特性的命中,具有合理的体外和体内PK特性,但不幸地被发现是磷酸二酯酶4的抑制剂。随后的药物化学项目优化了这个起点,用于口服PK,提高了GPR84效力,确认了NAM药理学和整体良好的选择性。GLPG1205进入了临床,虽然实现了适当的PK、安全性和耐受性特性,但在IBD的IIa期研究中没有实现主要疗效终点。在特发性肺纤维化患者中进一步评估了II期研究,并计划了III期研究,但Galapagos在2021年战略性地停止了该项目。
1.20.GPCR生物标志物
为了在临床前和临床试验中有效开发生物标志物,需要了解潜在疾病病理生理学和目标参与。生物标志物可以在临床前用于选择适当的模型和监测目标效应,而在临床上用于患者选择或临床研究的分层,或作为疾病进展的生物标志物,监测随时间的干预或疾病状态的变化,或监测药物安全性。因此,生物标志物可以监测潜在新药从实验室到临床的转化,并有助于通过反向转化回实验室更好地了解潜在的病理生理过程。
FDA将生物标志物定义为可以客观测量和评估的特征,作为正常生物过程、病理过程或对治疗干预的药理学反应的指标。这个定义允许任何有助于识别疾病机制和/或治疗效应的东西作为生物标志物,从代谢物、蛋白质或基因或蛋白质/基因模式(生物签名)、生物结构的变化、生物过程或特征特征。GPCR目标参与在体外通过信号过程的变化、蛋白质或基因表达或细胞激活状态很容易测量,如本文前面部分所述,所有这些都可以作为体外和体内的生物标志物,并且许多将提供从动物模型到人类的转化。
1.20.1.临床前和临床生物标志物
生物标志物应该是可访问的,可以准确测量,并且应该在不损害样本完整性的情况下为研究增加最小的复杂性。相对直接的临床前和临床研究中容易分析的生物流体,如血液、血浆或尿液,可以用来研究GPCR功能。例如,对精神病患者(如精神分裂症和抑郁症患者)的淋巴细胞分析表明GPCR表达与疾病之间存在相关性,突出了它们作为外周生物标志物的潜力。在临床研究中,由于确保多个地点之间一致的样本处理存在问题,对活细胞、信号机制、磷酸蛋白和代谢物的分析通常更具挑战性。
由于动物模型只模拟了人类疾病机制的某些方面,并且显示出菌株间变异性,因此在选择动物模型时必须谨慎,以确保所选模型模仿了正在研究的疾病方面,并且这与患者的病理生理反应有关。不同的动物模型可能需要用于疾病的不同方面,因此对疾病机制的了解不足或不足可能导致临床转化不佳。即使在动物疾病模型中有很好的证据表明阻断了机制,这些也不总是很好地转化为临床研究。例如,动物实验表明CCR5是一个很好的RA治疗候选药物,而随后使用CCR5拮抗剂的临床试验未能证明治疗效应。
一种用于监测体内或体外GPCR目标参与的方法是测量受体-配体结合。体外GPCR占据研究在早期CNS药物开发中具有实用性,为药物暴露(剂量、血浆浓度、CSF和大脑)与目标参与之间的关系提供了额外的数据。如果这些临床前数据表明一定程度的目标参与,例如,在相关动物模型中的受体占据,那么随后确定人类目标参与可以用来指导人类概念验证(PoC)阶段的药物开发剂量,第二阶段临床试验,从而加速化合物的临床开发。
PET(正电子发射断层扫描)是一种功能成像技术,可以用来证明新化学实体在药物管理后与感兴趣的GPCR相互作用。PET成像在CNS药物发现中特别有价值,由于人类大脑的不可达性,使用小分子放射性示踪剂对于评估大脑渗透、目标参与、大脑功能和神经病理学至关重要。这需要建立体内目标占据与药理效应之间的联系,并且随着PET技术的最新进展,它已成为药物发现中的重要工具。例如,5-HT2A受体涉及各种大脑疾病,如精神分裂症、抑郁症和阿尔茨海默症,5-HT2A PET示踪剂,最著名的[11C]MDL100907和[18F]altanserin,已经在临床上使用了数十年,用于量化人类大脑中的5-HT2A受体。尽管找到提供适当目标参与的合适配体存在挑战,但PET成像在CNS药物开发的PoC确定中发挥了关键作用。然而,PET技术不仅用于CNS药物发现,也已经应用于其他治疗领域。例如,全身混合PET成像,结合计算机断层扫描和/或磁共振,用于为特定目标或生物标志物开发新的放射性配体,使评估否则难以接触的肿瘤成为可能。
成像生物标志物在药物发现和开发中越来越重要,作为发现和临床开发之间的关键桥梁。
在临床中,生物标志物可以提供有价值的数据,以便在早期阶段的临床试验中做出关键的开发决策,通过确认生物学效应并允许早期评估新药的药理活性。在临床试验的剂量递增阶段应用适当的药效生物标志物,可以为靶向参与提供证据,并在不使患者暴露于不必要的毒性的情况下给出估计的最佳剂量。生物标志物也可以在患者中进行检查,以确认机制的调节,以确定对疾病的药理影响,尽管对生物标志物的积极影响并不总是导致有利的临床结果,正如前面讨论的CCR5拮抗剂的情况。
最近,已经从每项研究的单一生物标志物转向更以患者为中心的基于精准医学的生物标志物方法,考虑到患者的遗传状态。采用这种基于精准医学的生物标志物方法,允许为特定靶向治疗选择最合适的患者,并且正在使用不同的试验设计来识别和验证潜在的生物标志物,以帮助实现这一点。
1.20.2.未来的临床应用
对于GPCR生物学,从分子科学到强大的成像平台的技术进步,有望加速新生物标志物的发现,这将促进发现治疗疾病的新方法。例如,使用无偏的、假设生成方法的GPCR组学(例如,使用RNA测序对组织和细胞类型进行鉴定和量化GPCR表达),已经发现了以前未被认识的GPCR,这些GPCR有助于功能反应和病理生理学,并且可能是治疗靶标。人们意识到GPCR过度表达和突变导致异常激活的促癌途径诱导肿瘤进展,也使GPCR作为某些类型的癌症的生物标志物获得了更多的关注。来自患者衍生细胞和细胞系的研究,以及与正常细胞前体的比较,导致了这样一种观念,即肿瘤可能具有过度表达GPCR的“GPCR签名”,这些GPCR是肿瘤类型特异性的,并且可以作为生物标志物和/或治疗干预的目标。GPCR,特别是趋化因子受体,在肿瘤浸润性免疫细胞和肿瘤微环境中的细胞上高度表达,并且可能有助于特定的生物标志物签名。
更新的多组学(基因组学、转录组学、蛋白质组学、代谢组学、单细胞组学)和分析技术的进步,为在药物发现过程的每个阶段发现和验证多个与GPCR相关的生物标志物提供了激动人心的机会,并能够从微小样本(包括生物流体如血液、血浆、尿液、痰液、组织或灌洗液或活检样本)中提取高含量信息。例如,先进的质谱技术和下一代蛋白质组检测原理,如基于寡核苷酸的技术,通常需要微小量的样本,并且可以更容易地应用于大型患者队列,以识别作为治疗靶向候选物的因果蛋白。与更成熟的组学层(如转录组学)相比,蛋白质组学提供了直接测量蛋白质水平的好处,从而有助于在临床上转化为最常用的蛋白质生物标志物。
这些多组学生物标志物方法有可能为理解疾病过程、治疗干预和患者分层提供更个性化的生物标志物签名,并且来自多个来源的数据整合可能会显著增加对所获得测量值的价值和信心。然而,这一重大挑战需要适当的分析、整合和理解这些大数据集的工具和方法。来自不同来源和研究所的高含量和多因素组学数据以及来自具有多个混杂因素的不同患者群体的小样本数量,使得这种类型的数据分析特别具有挑战性,并且仍然存在对稳健和准确分析的未满足需求,以将许多这些潜在生物标志物转化为稳健的诊断和/或预后工具。人工智能和机器学习的最新进展可能会对这种类型的分析产生重大影响。
1.21.未来展望
我们对GPCR药物发现的理解持续增长,识别出新的和更好的方法来调节GPCR功能,成功地提供新的和改进的药物来治疗患者。GPCRs将仍然是一个重要的药物靶标类别,特别是当许多人预测大多数GPCRs尚未被药物化时。未来由GPCR组学技术和更高保真度的目标验证技术驱动的更深入的疾病知识将提高我们的能力,将未被药物化的GPCR调节与期望的治疗结果联系起来。新的技术平台GPCR生物技术公司(例如,Salipro 、Septerna、LeadXpro、Structure Tx)将继续出现,提供新的和流程来增加为新的GPCR机会找到化学起点的成功。
结构使能的GPCR项目将变得更加普遍,因为随着冷冻电镜的出现和计算化学力量的增加,我们生成和利用相关、天然和稳定的结构的能力得到了提高。值得注意的是,在药物发现项目开始时获得GPCR结构将提高预测配体-受体结合并加速新化学起点的药物化学优化的前瞻性能力。
不足为奇的是,难以药物化的GPCRs往往具有更大、不太可追踪的配体口袋,并且通常不太成功地采用传统的药物发现方法。然而,随着GPCR结构和功能理解的最新科学和技术进步,药物猎人正在转向天然GPCR复合物,正如在过去两年中对新检测平台(例如,Septerna、Domain Tx、OMass)的显著生物技术投资所强调的那样。特别是,新的GPCR结构信息的应用将继续扩大,以至于人工智能(AI)驱动的基于硅的分子相互作用预测将使基于硅的命中发现和优化在未来的GPCR药物发现中变得更加主流。
能够常规地识别小分子药物在天然GPCR信号复合物中的位置以及如何调节特定细胞功能及其之外的激动人心的前景,很可能成为现实。这种扩展GPCR检测设计的能力将识别和利用更理想的药物机制,超越传统的激动剂和拮抗剂,如别构调节剂和偏向信号药物,以及利用整个GPCR互作组。未来GPCR药物发现的其他激动人心的途径包括靶向细胞内GPCRs、孤受体GPCRs的去孤化、靶向GPCR降解和受体二聚体化抑制剂。
不足为奇的是,一些经过充分验证的GPCRs对小分子药物调节的抵抗力已成为生物药物模式和基于结构的流程(例如,Struct Tx)的焦点,如抗体、纳米抗体(Tectonic Tx)和内源性蛋白质-配体方法(Orion Biotechnology)。总的来说,生物药物正在成为药物发现中更可行和主流的方法,因此我们可以预期,对于特定的、高价值的GPCRs,生物药物方法将是提供新治疗的最佳方式。
我们希望这篇综述为您提供了对发现GPCRs新药的一些重要方面的最新见解,以及这个药物靶标家族所提供和挑战的激动人心的未来(图10,图11)。
识别微信二维码,添加生物制品圈小编,符合条件者即可加入
生物制品微信群!
请注明:姓名+研究方向!
版
权
声
明
本公众号所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,不希望被转载的媒体或个人可与我们联系(cbplib@163.com),我们将立即进行删除处理。所有文章仅代表作者观点,不代表本站立场。
细胞疗法抗体药物偶联物
100 项与 V-0219 相关的药物交易
登录后查看更多信息
研发状态
10 条进展最快的记录, 后查看更多信息
登录
| 适应症 | 最高研发状态 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|---|
| 2型糖尿病 | 临床前 | 西班牙 | 2022-03-29 | |
| 肥胖 | 临床前 | 西班牙 | 2022-03-29 |
登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
| 研究 | 分期 | 人群特征 | 评价人数 | 分组 | 结果 | 评价 | 发布日期 |
|---|
No Data | |||||||
登录后查看更多信息
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用