预约演示
更新于:2025-05-07
Chymopapain
糜蛋白酶
更新于:2025-05-07
概要
基本信息
在研机构 |
权益机构- |
最高研发阶段批准上市 |
首次获批日期 美国 (1959-07-08) |
最高研发阶段(中国)批准上市 |
特殊审评- |
登录后查看时间轴
结构/序列
Sequence Code 21649806

来源: *****
关联
2
项与 糜蛋白酶 相关的临床试验NCT06713733
Comparison of Clinical Efficacy of Submucosal Dexamethasone and Oral Trypsin-chymotrypsin for Reduction of Postoperative Sequelae After Lower Third Molar Surgery

CTRI/2024/01/061594
A Cross-Sectional Study to Assess The Real World Utilization of Oral Trypsin:Chymotrypsin Enzymes In Management of Injuries (ARMOR Study) - ARMOR

100 项与 糜蛋白酶 相关的临床结果
登录后查看更多信息
100 项与 糜蛋白酶 相关的转化医学
登录后查看更多信息
100 项与 糜蛋白酶 相关的专利(医药)
登录后查看更多信息
1,948
项与 糜蛋白酶 相关的文献(医药)2025-04-01Food Chemistry
Chemical composition and techno-functional properties of high-purity water-soluble keratein and its enzymatic hydrolysates
Article
作者: Sinkiewicz, Izabela ; Kusznierewicz, Barbara ; Giblin, Linda ; Sommer, Agata ; Taraszkiewicz, Antoni ; Staroszczyk, Hanna
2025-02-01The Journal of Pain
Backbone breakthroughs: How rodent models are shaping intervertebral disc disease treatment
Review
作者: Kumar, Hemant ; Bhombe, Komal ; Tripathi, Gyanoday
2025-02-01Journal of Food Science
Structures, antioxidant, and angiotensin I‐converting enzyme (ACE)‐inhibitory activities of peptides derived from protein hydrolysates of three phenolics‐rich legume genera
Article
作者: Chang, Sam K. C. ; Pechan, Tibor ; Zhang, Yan
100 项与 糜蛋白酶 相关的药物交易
登录后查看更多信息
研发状态
10 条最早获批的记录, 后查看更多信息
登录
| 适应症 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|
| 水肿 | 中国 | 1985-01-01 | |
| 炎症 | 中国 | 1985-01-01 | |
| 虹膜睫状体炎 | 中国 | 1985-01-01 | |
| 创伤和损伤 | 中国 | 1985-01-01 |
登录后查看更多信息
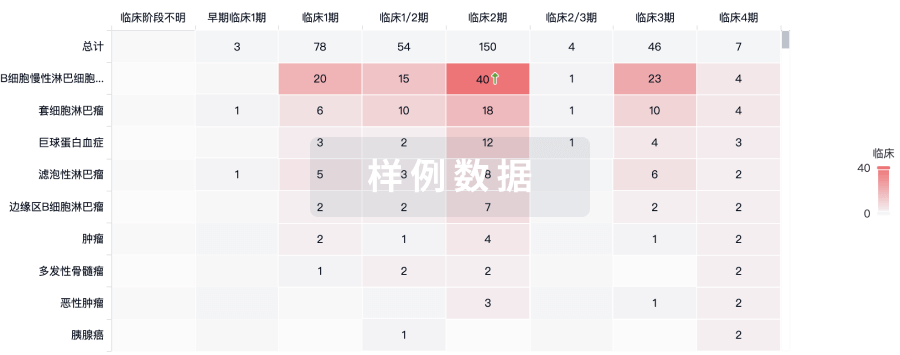
临床结果
临床结果
适应症
分期
评价
查看全部结果
登录后查看更多信息
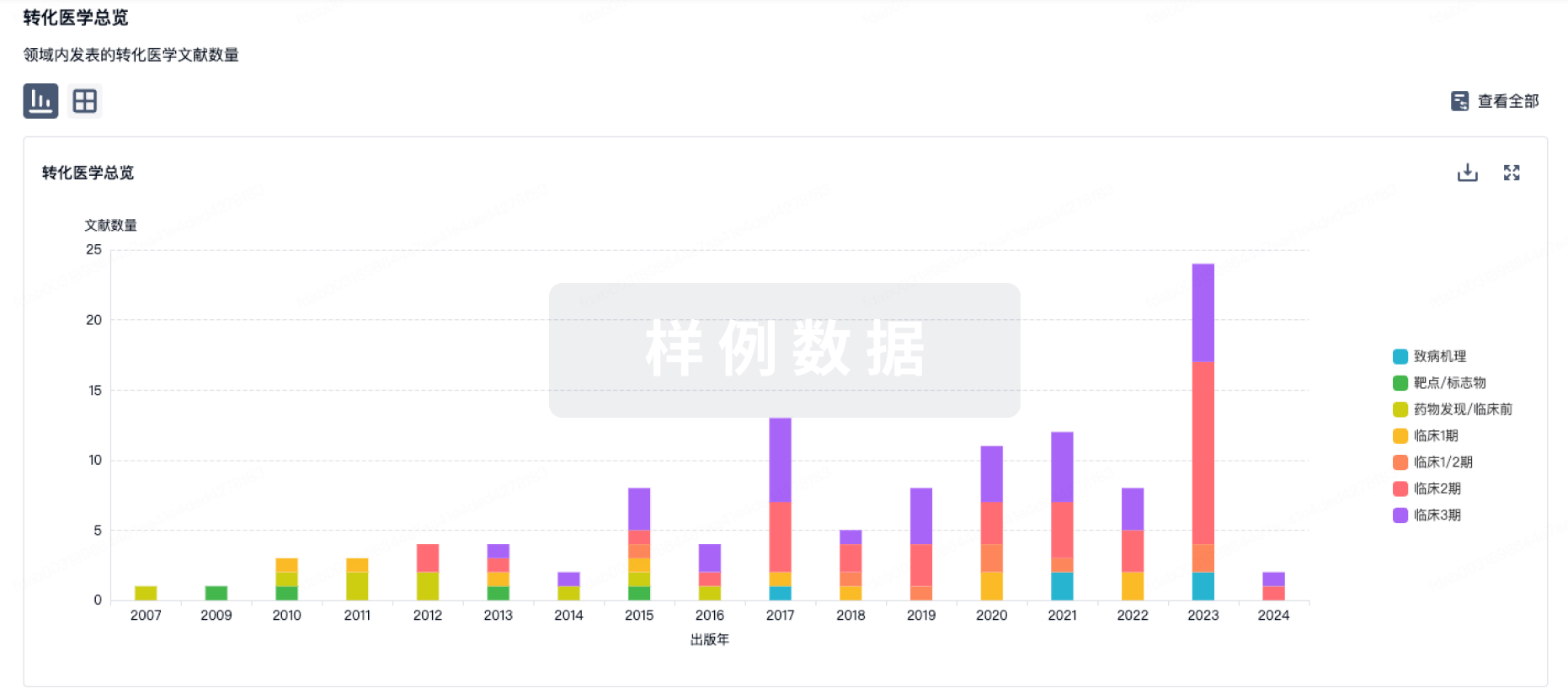
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
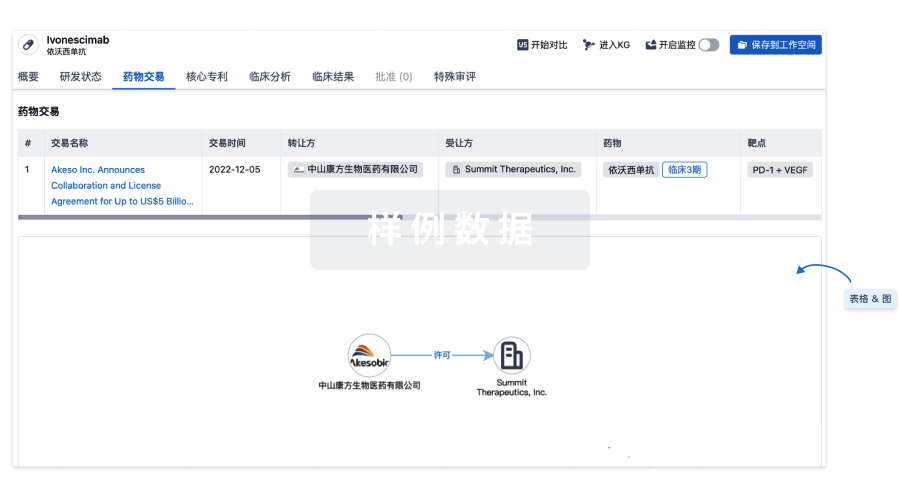
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
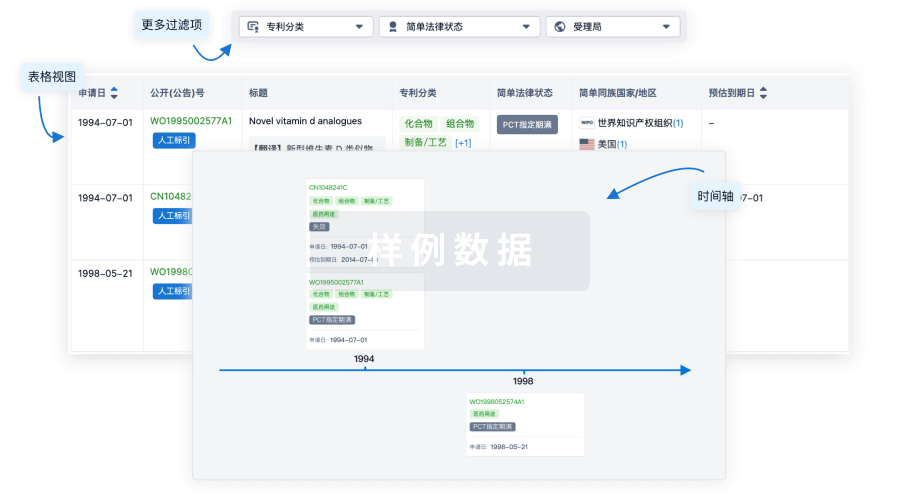
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
生物类似药
生物类似药在不同国家/地区的竞争态势。请注意临床1/2期并入临床2期,临床2/3期并入临床3期
登录
或
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
Eureka LS:
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用