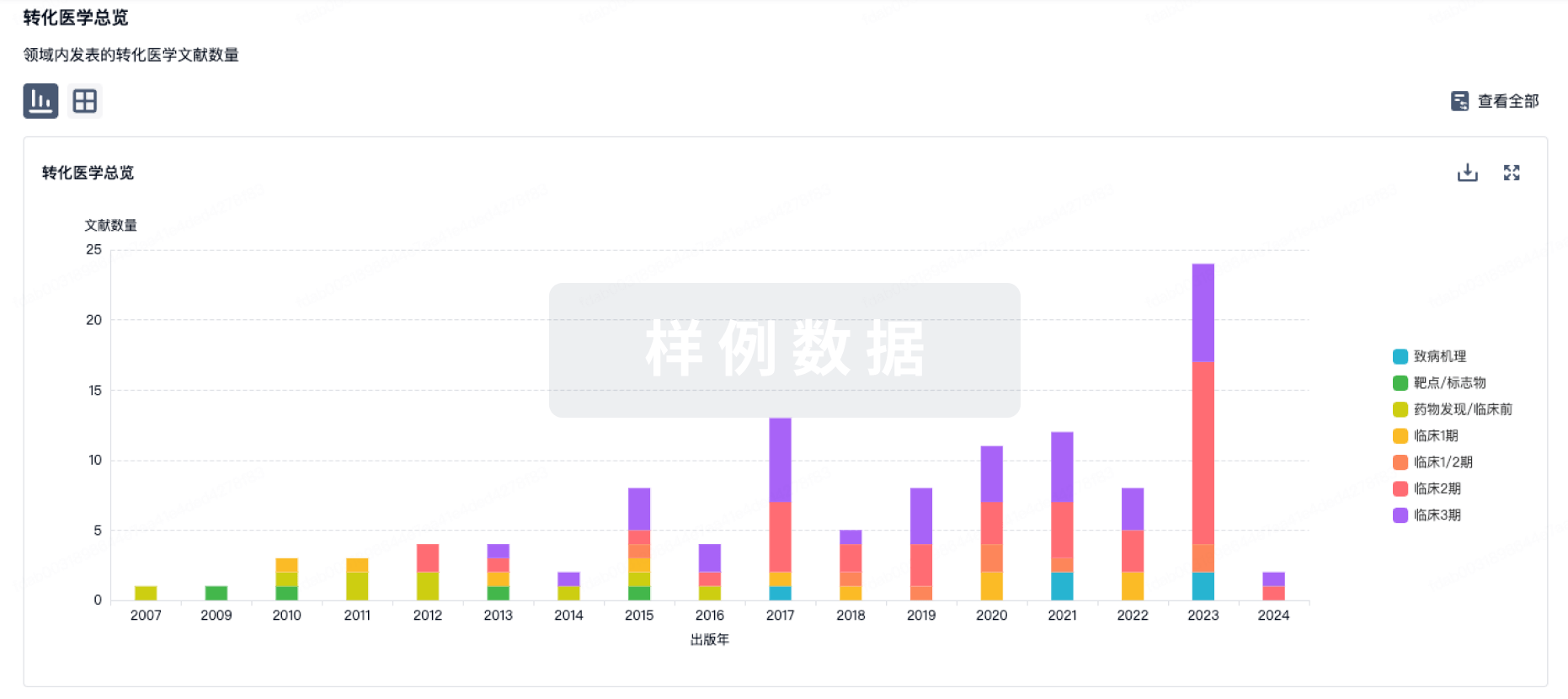
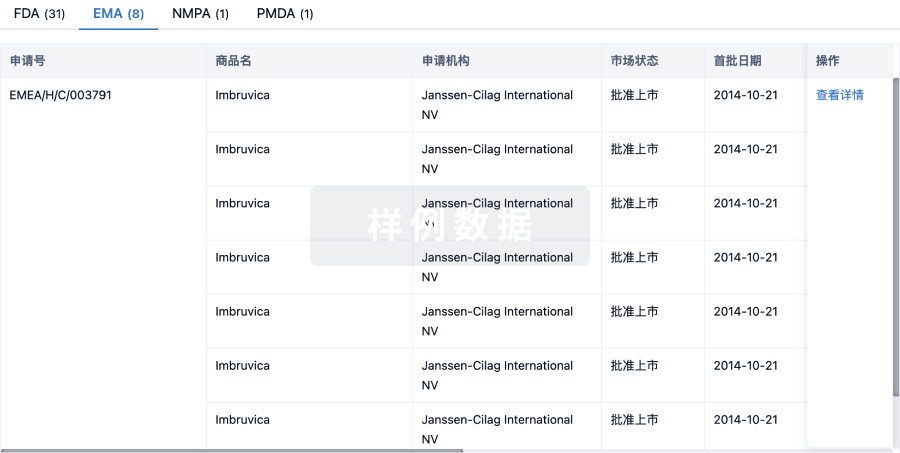
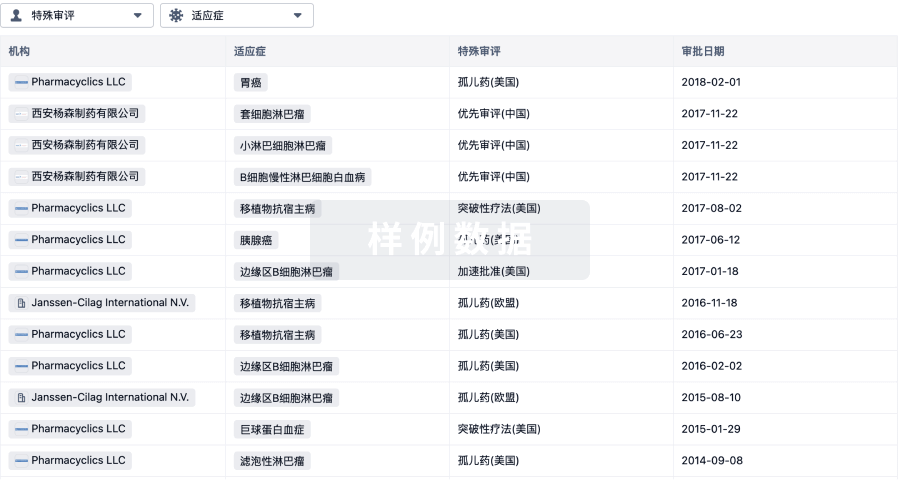
▲速抢免费参会名额 | 大湾区(广州)生物医药创新者峰会
KRAS 抑制剂的研究进展
来源
药学学报
作者
许俨钊,文辉,崔华清
中国医学科学院、北京协和医学院药物研究所, 活性物质发现与适药化研究北京市重点实验室
摘要
RAS(rat sarcoma)基因是一种重要的致癌基因,其突变存在于约30%的人类肿瘤中。KRAS(kirsten rat sarcoma viral oncogene)是RAS 的三个亚型之一,且相比于其他两种RAS 亚型更易出现突变。近年来,研究的不断深入为靶向KRAS治疗肿瘤提供了新的可能性,KRAS 抑制剂的研发也因此取得了显著的进展。本文根据各类抑制剂的作用模式,将KRAS 抑制剂分为直接靶向于KRAS 的抑制剂和间接作用于KRAS 的抑制剂两类,并由此综述介绍了具有代表性的KRAS抑制剂。
关键词
RAS;KRAS;作用模式;小分子抑制剂;抗肿瘤
_正文_
1982 年,研究者们[1-4]在人类膀胱癌细胞中发现了突变的RAS(rat sarcoma)基因,这使RAS 成为首个被发现的人类肿瘤基因[5, 6]。研究发现RAS 主要有HRAS、NRAS 和KRAS 三种亚型[7]。
RAS 在细胞信号转导过程中发挥核心作用[8]:在分子水平上可以调控转录、翻译等过程,在细胞水平上可以调节细胞增殖、分化、衰老、凋亡等等[9-11]。RAS的突变致癌约占人类癌症的30% [12],因此,RAS 一直被视为肿瘤治疗的重要靶点。
RAS 蛋白(图1)是一种低分子质量(21 kDa)、位于细胞膜上、拥有GTPase酶活性的鸟嘌呤核苷酸结合蛋白。RAS 主要由一个GTPase 催化结构域(残基1~166)和紧随其后的高度可变区域(highly variable region,HVR;残基167~188)组成[13]。各个亚型中GTPase 结构域的序列和结构几乎相同,而HVR 的序列存在显著差异[14, 15]。
RAS 通过在非活性状态(与GDP 结合)和活性状态(与GTP 结合)之间循环,发挥着二元开关的作用[13, 16]:在没有信号刺激时,这种循环十分缓慢。一旦接收到信号,RAS 就会在细胞膜上与被募集过来的鸟嘌呤核苷酸交换因子(guanine nucleotide-exchange factor;GEF)结合,释放GDP,并迅速与GTP 结合[17],进入“开”的阶段;随后在GTP 酶激活蛋白(GTPase-activating protein; GAP)的作用下,GTPase 的活性被大幅度增强[18],与RAS 结合的GTP 被水解为GDP,重新进入“关”的状态。绝大多数的RAS 突变会导致GTP 的水解受到抑制,RAS的活性无法被淬灭,进而引起RAS 的功能被异常激活。因此GTP 的水解成为了二元开关过程中的关键步骤。
根据RAS 的结构和调控机制,研究者们发现,在治疗RAS 方面存在种种难题[19, 20]:①RAS 蛋白上的GTPase 催化结构域高度保守,各个亚型之间很难做到高度选择性;②RAS 与GDP/GTP 的亲和力常数能达到皮摩尔级别,而细胞中的GTP 浓度是毫摩尔级别,因此小分子药物很难与底物进行竞争性结合;③RAS蛋白表面缺少有利于小分子结合的腔。
但是,随着新技术的发展,近年来针对RAS 的抑制剂又重新回到了人们的视野中。研究者们首先将目标放在了RAS 家族的亚型之一KRAS 上面。KRAS基因突变占RAS 基因突变总数的85%(NRAS 占12%,HRAS 占3%)[21]。在结肠癌、肺癌等多种人类癌症当中,经常能观察到KRAS 基因突变[22-24],而且在超过90% 的胰腺癌患者体内KRAS 基因都发生了突变[25]。因此,KRAS 成为了有效治疗RAS 的重要突破点。
1KRAS 抑制剂的研究进展
按抑制剂的作用模型将KRAS 抑制剂分为直接靶向KRAS 的抑制剂和间接作用于KRAS 的抑制剂两类。前者是直接以KRAS 为靶点的抑制剂,后者主要是通过抑制KRAS 信号通路的上下游部分,间接地对KRAS 起抑制作用。
1.1 直接靶向KRAS 的抑制剂
绝大多数的RAS 致癌突变集中在Gly12、Gly13、Gln61 这三个氨基酸残基上[12, 13, 26, 27]。其中,KRASG12C 突变占所有非小细胞肺癌(NSCLC)的病例超过59%,占所有KRAS 突变驱动的癌症的12%[12, 19, 28]。而基于半胱氨酸(Cys)共价结合抑制剂的成功研制,让KRASG12C 成为了治疗RAS 变异的理想靶点:在KRASG12C 中,12 号位的甘氨酸突变为半胱氨酸,而Cys 是一种易于形成共价键的氨基酸,如果小分子药物能够与这种Cys 结合,这类抑制剂将减少对结合腔的依赖,也可以获得相对较高的特异性。因此,特异性针对KRASG12C 的共价抑制剂,为靶向KRAS 药物的开发提供了一个重要的参考方向。
1.1.1 6H05 系列化合物
KRASG12C 共价抑制剂的发现是通过一项名为“tethering”的技术[19, 29]。“tethering”技术的基本原理是:针对含有Cys 的靶点,利用含双硫键的小分子碎片库进行筛选--小分子碎片的双硫键与KRASG12C 蛋白上Cys 的巯基发生交换后,小分子碎片以Cys 为“桥梁”与KRAS 蛋白相连接,小分子碎片在蛋白上找到合适的结合腔后便能附着下来,随后通过还原反应将作为“桥梁”的Cys 给清除掉。
通过“tethering”技术,在480 个化合物中筛选出了两个先导物,显示出对突变KRASG12C 的修饰程度最大,被命名为6H05 和2E07[29](图2)。随后通过对6H05 的构效关系进行研究,进一步优化确定了两个有效的化合物(化合物3 和化合物4,图2),这些化合物被称为6H05 系列[30]。
6H05 系列的化合物与KRASG12C 结合,发现了一个被称为Switch-Ⅱ pocket(S-ⅡP)的新结合位点[31]。晶体结构显示,S-ⅡP 位于KRAS 中央的β 折叠和Switch-Ⅱ (SW2)之间,是通过抑制剂结合并诱导SW2 重新排列而形成的[30]。结果证明6H05 系列让KRASG12C 共价抑制剂的尝试获得了首次成功。而S-ⅡP的发现意味着KRAS 蛋白中有了额外的结合位点,成为了后来药物设计的一个关键点。
1.1.2 喹唑啉系列化合物
KRASG12C 共价抑制剂发展进程中的另一个突破是喹唑啉系列化合物的发现。研究者[32]设计了一组喹唑啉化合物能够结合在S-IIP 空腔中,并在此基础上进一步的进行结构优化(图3)。结果表明,酰胺取代基的加入能够提高药物活性:除了增强抗增殖能力和诱导凋亡能力以外,酰胺基的修饰还增强了对KRASG12C 癌细胞的选择性。S-IIP 的发现以及喹唑啉结构的发现与优化,为后续KRASG12C 抑制剂的发展奠定了坚实的基础。
1.1.3 ARS 系列化合物
ARS 系列化合物是Wellspring 公司研发的一系列KRASG12C 抑制剂。其中比较有代表性的是ARS-853[33]和ARS-1620[34] (图4)。研究发现,KRASG12C 是ARS-853 在2 700 多种细胞蛋白质中最能有效进行共价结合的靶点[33]。ARS-853 与KRASG12C 的结合速率常数为76 L·mol -1·s-1,细胞结合IC50 在6 h 内为1.6 μmol·L-1。细胞实验表明,ARS-853 对多种肿瘤细胞系均具有抑制活性,表现出对肿瘤细胞凋亡的促进以及对肿瘤细胞生长的抑制。
ARS-853 仅与非活性状态的KRASG12C 结合,表明KRASG12C 不是停留在异常活跃的状态,而是在与GDP/GTP 结合态之间快速循环[33]。ARS-853 化合物虽然在低微摩尔范围内仍表现出较好的细胞活性,但存在致命的缺点:血浆代谢稳定性差(t1/2 < 0.2 h 人, t1/2 < 0.3 h 小鼠),而且小鼠口服生物利用度低(F < 2%)[34]。这导致ARS-853 无法用于进一步的体内研究。
Wellspring 公司基于喹唑啉的结构基础,对喹唑啉母核周围的取代基进行系统优化后,到得了几个活性较高的化合物,其中引人注目的是ARS-1620[34]。ARS-1620 是一种对KRASG12C 具有高效价和高选择性的共价化合物,与KRASG12C 的结合速率相比于ARS-853 提高了10 倍。细胞实验表明,在KRASG12C细胞系(H358、MIA-PaCa2、LU65)中,ARS-1620 的IC50 能达到150 nmol·L-1,在其他KRAS 突变的细胞系(H441、A549、HCT116)中也表现出良好的效果。在动物模型的进一步研究中,ARS-1620 在小鼠体内表现出极好的口服生物利用度(F > 60%)和足够的血液稳定性,而且在人源肿瘤异种移植瘤(patient-derivedtumor xenograft, PDX)模型中也证明了ARS-1620 对KRASG12C具有高度选择性,相比于ARS-853 有了显著的提高。实验数据也说明ARS-1620 可快速地实现体内靶标占据,抑制RAS 信号的传导,以此消灭掉肿瘤。
ARS-3248 是在ARS-1620 基础上研发的新一代KRASG12C 抑制剂,但其结构目前尚未被披露。目前,ARS-3248 正在开展临床I 期试验,选择KRASG12C 阳性晚期实体瘤患者入组。
1.1.4 MRTX849
MRTX849(图5)是由Mirati Therapeutics 公司开发的针对KRASG12C 的高度选择性的共价抑制剂,能够稳定地与处于非活性状态的KRASG12C 结合[35]。在100 mg·kg-1·d-1 的最大有效剂量下,MRTX849 针对不同的肿瘤模型均显示出剂量依赖性的抗肿瘤作用。相比于野生型KRAS 以及其他蛋白中的Cys 来说,MRTX849 针对KRASG12C 的选择性高了1 000 倍以上。MRTX849 口服生物利用度能达到30%,单次给药后半衰期长达25 h[30]。临床试验显示[36],MRTX849 临床试验招募了17 名患者,其中包括10 名NSCLC 患者、4 名结肠直肠癌(CRC)患者和3 名其他癌症类型患者。他们接受不同剂量的治疗,分别为150、300、600、1200 mg 每天口服一次,以及600 mg每天口服两次。在接受所有剂量治疗的患者中,3/6 的NSCLC 患者,以及1/4的CRC 患者能实现了部分缓解(partial response, PR);在接受最高剂量治疗的患者中,3/5 例的NSCLC 患者,和1/2 例的CRC 患者达到部分缓解,其他患者病情稳定(stable disease, SD)。
1.1.5 AMG 510
AMG 510(图6)是Amgen 公司开发的一种小分子抑制剂,也是第一个进入临床阶段的KRASG12C 共价抑制剂[37]。AMG 510 通过将KRAS 锁定在非活性的状态(与GDP 结合),特异性地抑制KRASG12C。
在肿瘤模型中,AMG 510 能选择性地靶向KRASG12C,并且单药治疗就可引起KRASG12C 肿瘤小鼠模型的肿瘤消退[38]。在临床试验中[39],AMG 510 依然表现出良好的耐受性,且没有剂量限制的毒性。在招募的34 位NSCLC 患者中,有23 位患者的疗效可被评估。其中有13 名患者接受了每天一次960 mg 目标剂量的治疗,其中7 名实现了部分缓解,6 名患者的病情稳定,控制率达到100%。
1.1.6 SML 系列
以GDP/GTP 结合位点进行共价激酶抑制剂的研发[40-42],为KRASG12C 抑制剂的研究提供了新的思路,得到了一系列核苷酸类的KRASG12C共价抑制剂。
基于GDP 的结构首先设计了一系列针对催化位点的、底物竞争相关的共价抑制剂,SML-8-73-1(图7)被确定为主要候选物[43]。模拟细胞条件,在1 mmol·L-1的GDP/GTP 存在的情况下,对SML-8-73-1 的结合效率进行测定,结果表明,在孵育2 h 后,SML-8-73-1 进行底物竞争性结合的KRASG12C 超过95%。
然而,SML-8-73-1 含有两个带负电荷的磷酸基团,导致难以穿过细胞膜。“caging”技术,也即合成各种修饰磷酸,如磷酸酯,是一种有效缓解甚至解决该问题的常见策略,它能通过化学修饰将带电离子部分所掩盖,从而使细胞能被动吸收[44]。在此基础上,对SML-8-73-1 的磷酸基团进行修饰,合成出了SM-10-70-1(图7)。SM-10-70-1 表现为细胞通透性增强,并通过共价结合竞争性地抑制KRASG12C。此外,KRAS 依赖的信号通路,如Akt 和ERK 通路也被抑制。不仅如此,在几个表达KRAS G12C 突变的癌细胞株中,证实了SM-10-70-1 具有抗增殖活性的能力。但其有效率和选择特异性仍有待进一步提高。因此,新的SARs研究得以继续,获得了有希望的化合物16[45(] 图7)。通过亚甲基取代SML-8-73-1和SM-10-70-1 磷酸酐键中的中心氧,化合物16 的化学稳定性和酶稳定性有了很大的提高。尽管相比于SML-8-73-1 降低了40 倍的亲和力,化合物16 仍然可能是一个有发展潜力的化合物。
1.2 间接作用于KRAS 的抑制剂
1.2.1 靶向KRAS 的mRNA 肿瘤疫苗
mRNA 肿瘤疫苗的治疗思路是:在肿瘤细胞发生基因突变后,会特异性表达新的抗原,通过对肿瘤细胞进行DNA 或RNA 测序,寻找出这类新抗原,随后以相对应的mRNA 为基础完成疫苗的制备,输回体内激活免疫细胞,进而杀灭带有上述新抗原的肿瘤细胞[46]。其中,构建的疫苗被称为mRNA 肿瘤疫苗。
有研究发现,从一种转基因小鼠体内分离并改造后的T 细胞,对多种类型肿瘤患者体内的KRAS 突变体均有识别作用[47],在后续实验中,有研究者[48]成功地从一名KRAS 突变的癌症患者身上移植培育出可被该T 细胞识别的细胞系。在此基础上,Moderna Therapeutics 公司与Merck & Co.合作开发了一种名为mRNA-5671 的mRNA 肿瘤疫苗,mRNA-5671 能在体内产生大量的抗原,引导T细胞寻找并攻击表达4 种关键KRAS 突变体(G12C、G12D、G12V 和G13C)的细胞[49]。mRNA-5671 可以直接在宿主细胞中表达KRAS 抗原,比基于DNA的疫苗更容易降解,减少了不良反应,比基于肽的疫苗也能更有效地促进T 细胞做出免疫反应[46, 50]。目前,为了更加充分地利用免疫系统能力,mRNA-5671在与PD1 抑制剂pembrolizumab 联合使用[49]。
1.2.2 SOS1 抑制剂
GEF 承担着介导KRAS 与GTP 结合的重任,因此,抑制GEF可以减少KRAS 与GTP 结合的机会,可能是靶向RAS 驱动肿瘤的一种可行方法[51]。而且,GEF 抑制剂类化合物阻断的是GEF 与KRAS 的相互作用,与KRAS是何种突变无太大关联。
SOS(son of sevenless)是常见的GEF 之一,包括SOS1 和SOS2 两种常见的亚型[52]。作为一种核苷酸交换因子,SOS 首先会与RAS 结合。RAS-SOS 复合物会改变RAS 蛋白的构象,改变后的构象与缺失镁辅因子(magnesium cofactor)的RAS(非活性状态)构象十分相似,因此阻断了镁辅因子与GDP 中的磷酸盐部分进行亲水性的相互作用[17],进而降低了RAS 与GDP 的亲和力,有助于GDP的释放。然后GTP 的鸟嘌呤核苷和核糖部分开始与RAS 结合,随后GTP 的γ-磷酸基团与RAS 的镁辅因子相互作用,导致RAS 的构象再次发生变化,SOS 从络合物中脱离出来。
由此可见,抑制SOS 的活性可降低RAS 蛋白对GTP 的结合能力,进而抑制RAS 的异常激活。基于这个思路,Boehringer Ingelheim(BI)公司找出了一些能够结合SOS1 并阻止SOS1 与RAS 相互作用的苗头化合物。随后BI 公司报道了BI-3406(IC50 = 6 nmol·L-1,图8)[53]。BI-3406 通过与SOS1 的催化位点结合,阻止KRAS-GDP 的相互作用。在单药治疗的情况下,BI-3406 能够有效地减少KRAS 依赖性的肿瘤细胞中活性形式(与GTP 结合)KRAS 的形成,并抑制MAPK 信号通路,引起肿瘤消退[53]。
随后BI 1701963 进入了临床开发阶段[54](目前未公布结构)。BI 1701963与BI-3406 密切相关,均能广泛针对KRAS 等位基因(G12D、G12V、G12C 以及G13D)表现出活性,同时保留了KRAS 与SOS2 的相互作用[55]。
此外,Bayer 公司也发现了一款SOS1 抑制剂[51],命名为BAY-293(IC50 = 21nmol·L-1,图9),可以选择性的抑制SOS1 活性,下调肿瘤细胞中RAS 的活性水平。BAY-293 在野生型KRAS 细胞中能够有效抑制RAS-RAF-MEK-ERK通路,且BAY-293 与ARS-853 的联用对于KRASG12C 细胞系的抗增殖活性表现出协同作用[51]。
1.2.3 二甲双胍
除了新药的研制,在“老药新用”方面,靶向KRAS 的研究也有重大突破:最近有研究团队发现,降糖一线药物二甲双胍(metformin,图10)也可用于靶向KRAS 的治疗,且二甲双胍的抑癌机制可能不依赖于其降糖作用[56]。研究发现,相比于其他降糖药,服用二甲双胍以后,携带KRAS 突变的转移性结直肠癌(metastatic colorectal cancer, mCRC)患者总生存时间延长了37.8 个月,然而二甲双胍不能提高野生型KRAS 患者的生存率。细胞实验发现,KRAS 的突变会导致膜通道蛋白MATE1 的表达水平下降,二甲双胍无法从肿瘤细胞中排出,进而在细胞中富集。上调癌细胞中MATE1 表达水平后二甲双胍的抑癌作用消失也证明了这一点。而高浓度的二甲双胍则通过抑制RAS/ERK 和AKT/mTOR 这两条通路,抑制了KRAS 突变癌细胞的增殖[56]。目前研究团队已经安排了前瞻性的临床研究。
1.2.4 PDEdelta 抑制剂
KRAS 与GTP 结合后会将效应蛋白分泌到质膜(plasmamembrane, PM)上,随后激活细胞中的增殖和存活信号[57, 58]。而PDEdelta(phosphodiesterase 6 delta)作为一种膜转运蛋白,通过相互作用控制KRAS 蛋白的转运[59, 60]。因此,干扰PDEdelta 与KRAS 的结合为靶向KRAS 突变癌症提供了一种新方法。
Zimmermann 等[61]首先研发得到了一种小分子抑制剂deltarasin(图11),deltarasin 通过结合PDEdelta 的异戊二烯结合腔,抑制了KRAS-PDEdelta 的相互作用,干扰KRAS 在PM 上的定位,进而抑制下游通路信号,减少癌细胞的增殖。然而当浓度高于9 μmol·L-1时,deltarasin 在所有实验用的细胞系中都表现出细胞毒性[61],并且在NSCLC 中deltarasin 会诱导自噬[62]。第二种PDEdelta 抑制剂deltazinone 1(图11)有效避免了细胞毒性的问题,且对PDEdelta 有较高的亲和力,但是在体内实验中deltazinone 1 会被快速代谢掉[63]。目前,仍有NHTD(图11)等PDEdelta 抑制剂处于临床前研究[64],并在考虑与联合治疗的策略。
2总结与展望
目前来看,针对G12C 突变的抑制剂仍是靶向KRAS 治疗肿瘤的主流方向。而且,针对KRASG12C 抑制剂临床表现不够完美的情况来看,不少研究者也采用了联合用药的策略[35, 38, 49, 65]。例如,在小鼠体内使用AMG 510 会引起促进发炎性的肿瘤微环境,采用AMG 510 和PD1 阻断剂pembrolizumab 能够有效缓解这个问题[38],并且对KRASG12D 肿瘤细胞也有一定的排斥作用。此外,研究者将AMG 510 与MEK 抑制剂、MAPK 抑制剂以及SHP2 抑制剂等多种抑制剂分别组合使用,均表现出不错的疗效[49];MRTX849 与HER2 抑制剂、CDK4 抑制剂以及SHP2 抑制剂等小分子抑制剂的组合[49],在增强MRTX849 的反应活性以及减弱耐药性方面,也具有良好的前景;BI 1701963 也在寻求与MEK 抑制剂的联合使用,以增强更好的疗效[54, 55]。
此外,蛋白降解靶向嵌合体(proteolysis targeting chimera, PROTAC)技术凭借其独有的优势,在治疗KRAS 方面也有着不错的应用[66, 67]。但是PROTAC 本身依然面临着一些难题,例如分子质量较大导致的药代动力学性质不佳以及口服生物利用度不高[68],可用于PROTAC 的泛素连接酶E3 过少(不到10 种)[68, 69]等问题。
总之,这些新的在研药物(表1)[49]不仅开拓了KRAS 治疗的新领域,为不同类型的KRAS 突变患者带来了更多的治疗方法,也为研究RAS 基因提供了有价值的方向,有助于将历史上“不可成药的”RAS 的相关药物引入临床。
无论是新技术的研发,还是新药物的发现,亦或是如二甲双胍在内的“老药新用”开辟的新天地,都表明研究者通过有策略地靶向RAS 治疗肿瘤取得了越来越好的临床结果。如同KRAS 抑制剂的研究正在如火如荼地展开一样,人们对于RAS 机制的研究也没有停止脚步。随着技术的发展,RAS 也会更加清晰地被展现在我们面前,我们也终将会攻克这个癌症领域里的难关。作者贡献:许俨钊撰写论文初稿;文辉、崔华清修改论文并定稿。利益冲突:本文无相关利益冲突。
参考文献
详见 药学学报
版权声明:本文转自凡默谷,如不希望被转载的媒体或个人可与我们联系,我们将立即删除
【关于药融圈】药融圈围绕我国生物医药产业链,针对生物医药大数据、技术和资本投资、药融园(产业园)等开展系列系统性工作,促进我国生物医药产业健康发展,完善产业链,共同面对全球合作和竞争。
点分享
点点赞点在看